 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8s8e | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
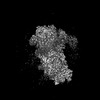
| タイトル | Structure of a yeast 48S-AUC preinitiation complex in closed conformation (model py48S-AUC-3.1) | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RIBOSOME / translation / initiation factors / 40S / eIF1A / AUC codon / eIF2 / tRNAi / 48S PIC / small ribosome subunit | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報formation of translation initiation ternary complex / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / eukaryotic translation initiation factor 3 complex, eIF3e / ABC-family proteins mediated transport / methionyl-initiator methionine tRNA binding / eukaryotic translation initiation factor 3 complex, eIF3m / incipient cellular bud site / translation reinitiation / eukaryotic translation initiation factor 2 complex ...formation of translation initiation ternary complex / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / eukaryotic translation initiation factor 3 complex, eIF3e / ABC-family proteins mediated transport / methionyl-initiator methionine tRNA binding / eukaryotic translation initiation factor 3 complex, eIF3m / incipient cellular bud site / translation reinitiation / eukaryotic translation initiation factor 2 complex / eukaryotic translation initiation factor 3 complex / formation of cytoplasmic translation initiation complex / multi-eIF complex / eukaryotic 43S preinitiation complex / formation of translation preinitiation complex / eukaryotic 48S preinitiation complex / positive regulation of translational fidelity / protein-synthesizing GTPase / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / Formation of a pool of free 40S subunits / L13a-mediated translational silencing of Ceruloplasmin expression / ribosomal small subunit binding / 90S preribosome / translation regulator activity / translation initiation factor binding / translation initiation factor activity / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / small-subunit processome / translational initiation / cytoplasmic stress granule / rRNA processing / double-stranded RNA binding / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytoplasmic translation / rRNA binding / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / GTPase activity / mRNA binding / protein kinase binding / GTP binding / nucleolus / RNA binding / zinc ion binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | |||||||||
| 生物種 |  Kluyveromyces lactis NRRL Y-1140 (酵母)Kluyveromyces lactis (酵母) Kluyveromyces lactis NRRL Y-1140 (酵母)Kluyveromyces lactis (酵母) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.85 Å | |||||||||
 データ登録者 データ登録者 | Villamayor-Belinchon, L. / Sharma, P. / Llacer, J.L. / Hussain, T. | |||||||||
| 資金援助 |  スペイン, スペイン,  インド, 2件 インド, 2件
| |||||||||
 引用 引用 | ジャーナル: Nucleic Acids Res / 年: 2024 タイトル: Structural basis of AUC codon discrimination during translation initiation in yeast. 著者: Laura Villamayor-Belinchón / Prafful Sharma / Yuliya Gordiyenko / Jose L Llácer / Tanweer Hussain /  要旨: In eukaryotic translation initiation, the 48S preinitiation complex (PIC) scans the 5' untranslated region of mRNAs to search for the cognate start codon (AUG) with assistance from various ...In eukaryotic translation initiation, the 48S preinitiation complex (PIC) scans the 5' untranslated region of mRNAs to search for the cognate start codon (AUG) with assistance from various eukaryotic initiation factors (eIFs). Cognate start codon recognition is precise, rejecting near-cognate codons with a single base difference. However, the structural basis of discrimination of near-cognate start codons was not known. We have captured multiple yeast 48S PICs with a near-cognate AUC codon at the P-site, revealing that the AUC codon induces instability in the codon-anticodon at the P-site, leading to a disordered N-terminal tail of eIF1A. Following eIF1 dissociation, the N-terminal domain of eIF5 fails to occupy the vacant eIF1 position, and eIF2β becomes flexible. Consequently, 48S with an AUC codon is less favourable for initiation. Furthermore, we observe hitherto unreported metastable states of the eIF2-GTP-Met-tRNAMet ternary complex, where the eIF2β helix-turn-helix domain may facilitate eIF5 association by preventing eIF1 rebinding to 48S PIC. Finally, a swivelled head conformation of 48S PIC appears crucial for discriminating incorrect and selection of the correct codon-anticodon pair during translation initiation. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8s8e.cif.gz | 2.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8s8e.ent.gz | 1.6 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8s8e.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8s8e_validation.pdf.gz | 1.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8s8e_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 8s8e_validation.xml.gz | 192.4 KB | 表示 | |
| CIF形式データ | 8s8e_validation.cif.gz | 318.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/s8/8s8eftp://data.pdbj.org/pub/pdb/validation_reports/s8/8s8e | HTTPS FTP |
-関連構造データ
| 関連構造データ |  19802MC  8rw1C  8s8dC  8s8fC  8s8gC 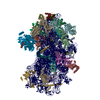 8s8hC 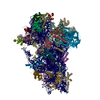 8s8iC  8s8jC  8s8kC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |





-リンク
 PDBj
PDBj
































- 集合体
集合体
| 登録構造単位 | 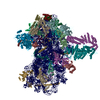
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 3種, 3分子 231
| #1: RNA鎖 | 分子量: 579454.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) |
|---|---|
| #37: RNA鎖 | 分子量: 15304.940 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Kluyveromyces lactis (酵母) |
| #38: RNA鎖 | 分子量: 24713.969 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) |
-Small ribosomal subunit protein ... , 11種, 11分子 ABCGOWQUcdf
| #2: タンパク質 | 分子量: 28264.525 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CN12 |
|---|---|
| #3: タンパク質 | 分子量: 28971.643 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CWD0 |
| #4: タンパク質 | 分子量: 27649.979 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CKL3 |
| #6: タンパク質 | 分子量: 26970.391 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CM04 |
| #12: タンパク質 | 分子量: 14530.655 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: P27069 |
| #14: タンパク質 | 分子量: 14645.041 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CW21 |
| #26: タンパク質 | 分子量: 15874.531 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q875N2 |
| #30: タンパク質 | 分子量: 13337.604 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CIM1 |
| #32: タンパク質 | 分子量: 7549.824 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: P33285 |
| #33: タンパク質 | 分子量: 6662.570 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CPG3 |
| #34: タンパク質 | 分子量: 17110.977 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: P69061 |
-40S ribosomal protein ... , 12種, 12分子 EHIVYabehDMZ
| #5: タンパク質 | 分子量: 29617.514 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CWJ2 |
|---|---|
| #7: タンパク質 | 分子量: 21735.297 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CTD6 |
| #8: タンパク質 | 分子量: 22642.727 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CMG3 |
| #13: タンパク質 | 分子量: 9797.949 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CXT6 |
| #16: タンパク質 | 分子量: 15194.549 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CU44 |
| #17: タンパク質 | 分子量: 13539.957 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CS01 |
| #18: タンパク質 | 分子量: 8884.362 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CNL2 |
| #19: タンパク質 | 分子量: 7141.421 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CUH5 |
| #20: タンパク質・ペプチド | 分子量: 3354.243 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) |
| #21: タンパク質 | 分子量: 26300.535 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CRK7 |
| #24: タンパク質 | 分子量: 14466.398 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CLU4 |
| #31: タンパク質 | 分子量: 12002.116 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CW78 |
-タンパク質 , 12種, 12分子 JLNXFKPRSTgi
| #9: タンパク質 | 分子量: 21587.049 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CM18 |
|---|---|
| #10: タンパク質 | 分子量: 17843.930 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CX80 |
| #11: タンパク質 | 分子量: 16989.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CJK0 |
| #15: タンパク質 | 分子量: 16047.897 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: F2Z602 |
| #22: タンパク質 | 分子量: 25385.975 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CRA3 |
| #23: タンパク質 | 分子量: 12584.377 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CVZ5 |
| #25: タンパク質 | 分子量: 15986.796 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CKV4 |
| #27: タンパク質 | 分子量: 15722.216 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CWU3 |
| #28: タンパク質 | 分子量: 17084.602 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CWT9 |
| #29: タンパク質 | 分子量: 15879.010 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CXM0 |
| #35: タンパク質 | 分子量: 35830.945 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis NRRL Y-1140 (酵母) / 参照: UniProt: Q6CNI7 |
| #36: タンパク質 | 分子量: 17462.168 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: TIF11, YMR260C, YM8156.02C / 発現宿主:  |
-Eukaryotic translation initiation factor 2 subunit ... , 3種, 3分子 jkl
| #39: タンパク質 | 分子量: 34763.652 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SUI2, TIF211, YJR007W, J1429 / 発現宿主: |
|---|---|
| #40: タンパク質 | 分子量: 57942.699 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: GCD11, SUI4, TIF213, YER025W / 発現宿主: |
| #41: タンパク質 | 分子量: 31631.309 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SUI3, TIF212, YPL237W / 発現宿主: |
-Eukaryotic translation initiation factor 3 subunit ... , 2種, 2分子 op
| #42: タンパク質 | 分子量: 110517.641 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: RPG1, TIF32, YBR079C, YBR0734 / 発現宿主: |
|---|---|
| #43: タンパク質 | 分子量: 93310.172 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: NIP1, YMR309C, YM9924.01C, YM9952.11C / 発現宿主: |
-非ポリマー , 5種, 124分子 


| #44: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 116 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 116 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#45: 化合物 |  分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION#46: 化合物 | ChemComp-GCP / |  分子量: 521.208 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C11H18N5O13P3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 521.208 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C11H18N5O13P3 / タイプ: SUBJECT OF INVESTIGATIONコメント: GMP-PCP, エネルギー貯蔵分子類似体*YM #47: 化合物 | ChemComp-MET / |  タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO2S / タイプ: SUBJECT OF INVESTIGATION タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO2S / タイプ: SUBJECT OF INVESTIGATION#48: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 3 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 1.8 MDa / 実験値: NO | ||||||||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 6.5 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.1 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 59000 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2.7 mm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最高温度: 100 K / 最低温度: 90 K |
| 撮影 | 平均露光時間: 1.1 sec. / 電子線照射量: 30 e/Å2 / 検出モード: INTEGRATING フィルム・検出器のモデル: FEI FALCON III (4k x 4k) 撮影したグリッド数: 8 / 実像数: 11245 詳細: Images were collected in movie-mode at 30 frames per second |
- 解析
解析
| EMソフトウェア | 名称: PHENIX / バージョン: 1.20.1_4487: / カテゴリ: モデル精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 画像処理 | 詳細: FEI Falcon III | ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1680000 / 詳細: Selected particles after 2d-classification | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.85 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 34495 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: OTHER / 空間: RECIPROCAL / Target criteria: FSC | ||||||||||||||||||||||||
| 拘束条件 |
|
