 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8p85 | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | 80S yeast ribosome in complex with Fluorolissoclimide | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / eL42 / Translation inhibitors | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報triplex DNA binding / ribosome hibernation / translation elongation factor binding / Platelet degranulation / regulation of translational initiation in response to stress / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity ...triplex DNA binding / ribosome hibernation / translation elongation factor binding / Platelet degranulation / regulation of translational initiation in response to stress / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation / mTORC1-mediated signalling / Protein hydroxylation / ribosome-associated ubiquitin-dependent protein catabolic process / GDP-dissociation inhibitor activity / regulation of polysaccharide biosynthetic process / positive regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / preribosome, small subunit precursor / nonfunctional rRNA decay / transporter complex / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / response to cycloheximide / telomeric DNA binding / Major pathway of rRNA processing in the nucleolus and cytosol / mRNA destabilization / lipopolysaccharide transport / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / negative regulation of translational frameshifting / TOR signaling / Formation of a pool of free 40S subunits / positive regulation of protein kinase activity / preribosome, large subunit precursor / L13a-mediated translational silencing of Ceruloplasmin expression / Gram-negative-bacterium-type cell outer membrane assembly / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translational elongation / ribosomal large subunit export from nucleus / G-protein alpha-subunit binding / 90S preribosome / Ub-specific processing proteases / ribosomal subunit export from nucleus / regulation of translational fidelity / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / protein-RNA complex assembly / translational termination / maturation of LSU-rRNA / ribosomal small subunit export from nucleus / translation regulator activity / translation repressor activity / DNA-(apurinic or apyrimidinic site) endonuclease activity / rescue of stalled ribosome / telomere maintenance / protein kinase C binding / cellular response to amino acid starvation / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosomal large subunit biogenesis / ribosome assembly / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / macroautophagy / small-subunit processome / translational initiation / modification-dependent protein catabolic process / protein tag activity / maintenance of translational fidelity / cell outer membrane / cytoplasmic stress granule / rRNA processing / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / 5S rRNA binding / small ribosomal subunit rRNA binding / ribosomal large subunit assembly / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / protein ubiquitination / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / G protein-coupled receptor signaling pathway / negative regulation of gene expression / response to antibiotic / mRNA binding / ubiquitin protein ligase binding / negative regulation of apoptotic process / nucleolus 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.9 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.9 Å | ||||||
 データ登録者 データ登録者 | Terrosu, S. / Yusupov, M. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: To Be Published タイトル: Role of Protein eL42 in the Ribosome Binding of Synthetic Analogues of Natural Lissoclimides 著者: Terrosu, S. / Yusupov, M. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8p85.cif.gz | 10 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8p85.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 8p85.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8p85_validation.pdf.gz | 3.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8p85_full_validation.pdf.gz | 4.8 MB | 表示 | |
| XML形式データ | 8p85_validation.xml.gz | 1.3 MB | 表示 | |
| CIF形式データ | 8p85_validation.cif.gz | 1.8 MB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/p8/8p85ftp://data.pdbj.org/pub/pdb/validation_reports/p8/8p85 | HTTPS FTP |
-関連構造データ
| 関連構造データ | |
|---|---|
| 類似構造データ |

-リンク
 PDBj
PDBj




























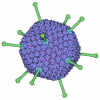




- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 単位格子 |
|
-要素
-RNA鎖 , 4種, 8分子 AsR1AR3AS4AT
| #1: RNA鎖 | 分子量: 579761.938 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #35: RNA鎖 | 分子量: 1097493.875 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #36: RNA鎖 | 分子量: 38951.105 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #37: RNA鎖 | 分子量: 50682.922 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
+40S ribosomal protein ... , 25種, 50分子 Bs0Cs1Ds2Fs4Gs5Hs6Is7Js8Ks9Mc1Oc3Pc4Qc5Rc6Sc7...
-Small ribosomal subunit protein ... , 5種, 10分子 Es3Lc0Vd0bd6ed9
| #5: タンパク質 | 分子量: 26542.789 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #12: タンパク質 | 分子量: 12757.445 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #21: タンパク質 | 分子量: 13929.044 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #73: タンパク質 | 分子量: 13480.886 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #76: タンパク質 | 分子量: 6675.723 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
+60S ribosomal protein ... , 38種, 76分子 AADBDA9ABDCACDDADDECDjAEDFCEkAFDGlCFmCGnCHoCIpCJqCK...
-Large ribosomal subunit protein ... , 2種, 4分子 sCMAODP
| #45: タンパク質 | 分子量: 19785.719 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #67: タンパク質・ペプチド | 分子量: 3354.243 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-タンパク質 , 5種, 9分子 ANDOisMp0ge1hRb
| #66: タンパク質 | 分子量: 14583.077 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #70: タンパク質 | 分子量: 30048.010 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #71: タンパク質 | | 分子量: 33749.121 Da / 分子数: 1 / 由来タイプ: 天然 詳細: There is something wrong in the extracted sequence, it does not match the one present in the pdb file. The first part actually matches, in the second part I modeled amminoacids from 189 till ...詳細: There is something wrong in the extracted sequence, it does not match the one present in the pdb file. The first part actually matches, in the second part I modeled amminoacids from 189 till the end. I do not understand what is going on with this particular chain. 由来: (天然) #78: タンパク質 | 分子量: 17254.227 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #79: タンパク質 | 分子量: 34841.219 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-非ポリマー , 8種, 2766分子 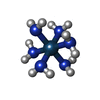


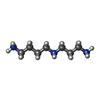



| #80: 化合物 | ChemComp-OHX /  分子量: 286.365 Da / 分子数: 1003 / 由来タイプ: 合成 / 式: H12N6Os 分子量: 286.365 Da / 分子数: 1003 / 由来タイプ: 合成 / 式: H12N6Os#81: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 1502 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1502 / 由来タイプ: 合成 / 式: Mg#82: 化合物 | ChemComp-K /  分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K 分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K#83: 化合物 |  分子量: 145.246 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C7H19N3 分子量: 145.246 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C7H19N3#84: 化合物 | 分子量: 367.455 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C20H30FNO4 / タイプ: SUBJECT OF INVESTIGATION #85: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 17 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 17 / 由来タイプ: 合成 / 式: Zn#86: 化合物 | ChemComp-5XU / (  タイプ: L-peptide linking / 分子量: 73.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H7NO タイプ: L-peptide linking / 分子量: 73.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H7NO#87: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 231 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 3.21 Å3/Da / 溶媒含有率: 61.73 % |
|---|---|
| 結晶化 | 温度: 277.15 K / 手法: 蒸気拡散法, ハンギングドロップ法 詳細: ll lissoclimides/80S complexes were formed in 5.5 mM Tris-acetate at pH 7.0, 3 mM K(OAc) at pH 7.2, 5.5 mM NH4(OAc), 2 mM Mg(OAc)2, 1.3 mM DTT by incubation of 80S ribosomes (1.5 uM) with 30- ...詳細: ll lissoclimides/80S complexes were formed in 5.5 mM Tris-acetate at pH 7.0, 3 mM K(OAc) at pH 7.2, 5.5 mM NH4(OAc), 2 mM Mg(OAc)2, 1.3 mM DTT by incubation of 80S ribosomes (1.5 uM) with 30-fold molar excess of lissoclimide congeners for 15 min at 30 C. Crystals were grown at 4 C by hanging-drop vapor diffusion |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: SLS  / ビームライン: X06SA / 波長: 1 Å / ビームライン: X06SA / 波長: 1 Å |
| 検出器 | タイプ: DECTRIS EIGER X 16M / 検出器: PIXEL / 日付: 2021年6月10日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 1 Å / 相対比: 1 |
| 反射 | 解像度: 2.9→207.6 Å / Num. obs: 3210225 / % possible obs: 99.69 % / 冗長度: 12 % / Biso Wilson estimate: 76.98 Å2 / CC1/2: 0.994 / Rmerge(I) obs: 0.09 / Rpim(I) all: 0.094 / Rrim(I) all: 0.13 / Net I/σ(I): 25.4 |
| 反射 シェル | 解像度: 2.9→3 Å / Mean I/σ(I) obs: 0.8 / Num. unique obs: 229131 / CC1/2: 0.12 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 / 解像度: 2.9→207.6 Å / 交差検証法: FREE R-VALUE
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.9→207.6 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
|
