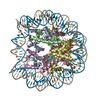
Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8of4 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Nucleosome Bound human SIRT6 (Composite) | ||||||
 Components Components |
| ||||||
 Keywords Keywords | GENE REGULATION / Transferase / Deacetylase / Histone H3 deacetylation | ||||||
| Function / homology |  Function and homology information Function and homology informationhistone H3K56 deacetylase activity, NAD-dependent / ketone biosynthetic process / histone H3K18 deacetylase activity, NAD-dependent / histone H3K9 deacetylase activity, hydrolytic mechanism / histone H3K9 deacetylase activity, NAD-dependent / protein delipidation / NAD+-protein-lysine ADP-ribosyltransferase activity / chromosome, subtelomeric region / regulation of lipid catabolic process / positive regulation of protein localization to chromatin ...histone H3K56 deacetylase activity, NAD-dependent / ketone biosynthetic process / histone H3K18 deacetylase activity, NAD-dependent / histone H3K9 deacetylase activity, hydrolytic mechanism / histone H3K9 deacetylase activity, NAD-dependent / protein delipidation / NAD+-protein-lysine ADP-ribosyltransferase activity / chromosome, subtelomeric region / regulation of lipid catabolic process / positive regulation of protein localization to chromatin / NAD+-protein-arginine ADP-ribosyltransferase activity / NAD-dependent protein demyristoylase activity / NAD-dependent protein depalmitoylase activity / DNA damage sensor activity / positive regulation of stem cell differentiation / negative regulation of D-glucose import across plasma membrane / positive regulation of chondrocyte proliferation / transposable element silencing / protein acetyllysine N-acetyltransferase / protein deacetylation / NAD-dependent protein lysine deacetylase activity / pericentric heterochromatin formation / histone deacetylase activity, NAD-dependent / positive regulation of blood vessel branching / cardiac muscle cell differentiation / protein localization to site of double-strand break / positive regulation of vascular endothelial cell proliferation / negative regulation of glycolytic process / TORC2 complex binding / negative regulation of protein import into nucleus / regulation of protein secretion / positive regulation of stem cell proliferation / regulation of double-strand break repair via homologous recombination / negative regulation of gene expression, epigenetic / positive regulation of double-strand break repair / lncRNA binding / NAD+-protein mono-ADP-ribosyltransferase activity / DNA repair-dependent chromatin remodeling / positive regulation of stem cell population maintenance / positive regulation of telomere maintenance / negative regulation of cellular senescence / regulation of lipid metabolic process / Transferases; Glycosyltransferases; Pentosyltransferases / negative regulation of transcription elongation by RNA polymerase II / NAD+ poly-ADP-ribosyltransferase activity / NAD+ binding / negative regulation of gluconeogenesis / positive regulation of fat cell differentiation / subtelomeric heterochromatin formation / pericentric heterochromatin / response to UV / nucleosome binding / regulation of protein localization to plasma membrane / site of DNA damage / negative regulation of protein localization to chromatin / nucleotidyltransferase activity / Transferases; Acyltransferases; Transferring groups other than aminoacyl groups / positive regulation of protein export from nucleus / determination of adult lifespan / circadian regulation of gene expression / chromatin DNA binding / regulation of circadian rhythm / base-excision repair / protein destabilization / Pre-NOTCH Transcription and Translation / positive regulation of fibroblast proliferation / protein import into nucleus / positive regulation of insulin secretion / structural constituent of chromatin / transcription corepressor activity / nucleosome / double-strand break repair / heterochromatin formation / nucleosome assembly / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / glucose homeostasis / positive regulation of cold-induced thermogenesis / site of double-strand break / Processing of DNA double-strand break ends / damaged DNA binding / chromatin remodeling / protein heterodimerization activity / negative regulation of cell population proliferation / chromatin binding / chromatin / negative regulation of transcription by RNA polymerase II / endoplasmic reticulum / protein homodimerization activity / DNA binding / zinc ion binding / nucleoplasm / nucleus Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.94 Å | ||||||
 Authors Authors | Smirnova, E. / Bignon, E. / Schultz, P. / Papai, G. / Ben-Shem, A. | ||||||
| Funding support |  France, 1items France, 1items
| ||||||
 Citation Citation | Journal: Elife / Year: 2024 Title: Binding to nucleosome poises human SIRT6 for histone H3 deacetylation. Authors: Ekaterina Smirnova / Emmanuelle Bignon / Patrick Schultz / Gabor Papai / Adam Ben Shem / Abstract: Sirtuin 6 (SIRT6) is an NAD-dependent histone H3 deacetylase that is prominently found associated with chromatin, attenuates transcriptionally active promoters and regulates DNA repair, metabolic ...Sirtuin 6 (SIRT6) is an NAD-dependent histone H3 deacetylase that is prominently found associated with chromatin, attenuates transcriptionally active promoters and regulates DNA repair, metabolic homeostasis and lifespan. Unlike other sirtuins, it has low affinity to free histone tails but demonstrates strong binding to nucleosomes. It is poorly understood how SIRT6 docking on nucleosomes stimulates its histone deacetylation activity. Here, we present the structure of human SIRT6 bound to a nucleosome determined by cryogenic electron microscopy. The zinc finger domain of SIRT6 associates tightly with the acidic patch of the nucleosome through multiple arginine anchors. The Rossmann fold domain binds to the terminus of the looser DNA half of the nucleosome, detaching two turns of the DNA from the histone octamer and placing the NAD binding pocket close to the DNA exit site. This domain shows flexibility with respect to the fixed zinc finger and moves with, but also relative to, the unwrapped DNA terminus. We apply molecular dynamics simulations of the histone tails in the nucleosome to show that in this mode of interaction, the active site of SIRT6 is perfectly poised to catalyze deacetylation of the H3 histone tail and that the partial unwrapping of the DNA allows even lysines close to the H3 core to reach the enzyme. #1: Journal: Elife / Year: 2023Title: Binding to nucleosome poises SIRT6 for histone H3 de-acetylation Authors: Smirnova, E. / Bignon, E. / Schultz, P. / Papai, G. / Ben-Shem, A. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8of4.cif.gz | 358 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8of4.ent.gz | 265.8 KB | Display | PDB format |
| PDBx/mmJSON format | 8of4.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/of/8of4ftp://data.pdbj.org/pub/pdb/validation_reports/of/8of4 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  16845MC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj








































- Assembly
Assembly
| Deposited unit | 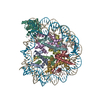
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 9 molecules AEBFCGDHL
| #1: Protein | Mass: 15421.101 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  #2: Protein | Mass: 11394.426 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #3: Protein | Mass: 14093.436 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #4: Protein | Mass: 13979.291 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #7: Protein | | Mass: 39180.914 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: SIRT6, SIR2L6 / Production host: References: UniProt: Q8N6T7, Transferases; Acyltransferases; Transferring groups other than aminoacyl groups, protein acetyllysine N-acetyltransferase, Transferases; Glycosyltransferases; Pentosyltransferases |
|---|
-DNA chain , 2 types, 2 molecules IJ
| #5: DNA chain | Mass: 44520.383 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|---|
| #6: DNA chain | Mass: 44991.660 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
-Non-polymers , 1 types, 1 molecules 
| #8: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION |
|---|
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight |
| ||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||
| Specimen support | Grid material: COPPER/RHODIUM / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R3.5/1 | ||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM imaging | Accelerating voltage: 300 kV / C2 aperture diameter: 50 µm / Cryogen: NITROGEN / Electron source:
| ||||||||||||
| Image recording |
| ||||||||||||
| EM imaging optics |
|
 FIELD EMISSION GUN
FIELD EMISSION GUN- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 2033169 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.94 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 439796 / Algorithm: BACK PROJECTION / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building |
|