response to mineralocorticoid / GMP binding / negative regulation of epithelial cell differentiation / forebrain astrocyte development / LRR domain binding / epithelial tube branching involved in lung morphogenesis / regulation of synaptic transmission, GABAergic / response to isolation stress / response to gravity / type I pneumocyte differentiation ...response to mineralocorticoid / GMP binding / negative regulation of epithelial cell differentiation / forebrain astrocyte development / LRR domain binding / epithelial tube branching involved in lung morphogenesis / regulation of synaptic transmission, GABAergic / response to isolation stress / response to gravity / type I pneumocyte differentiation / skeletal muscle cell differentiation / myoblast proliferation / Rac protein signal transduction / Signaling by RAS GAP mutants / Signaling by RAS GTPase mutants / Activation of RAS in B cells / cardiac muscle cell proliferation / RAS signaling downstream of NF1 loss-of-function variants / positive regulation of glial cell proliferation / RUNX3 regulates p14-ARF / SOS-mediated signalling / Activated NTRK3 signals through RAS / Activated NTRK2 signals through RAS / SHC1 events in ERBB4 signaling / Signalling to RAS / homeostasis of number of cells within a tissue / glial cell proliferation / SHC-related events triggered by IGF1R / Activated NTRK2 signals through FRS2 and FRS3 / Estrogen-stimulated signaling through PRKCZ / positive regulation of Rac protein signal transduction / SHC-mediated cascade:FGFR3 / MET activates RAS signaling / SHC-mediated cascade:FGFR2 / SHC-mediated cascade:FGFR4 / Signaling by PDGFRA transmembrane, juxtamembrane and kinase domain mutants / Signaling by PDGFRA extracellular domain mutants / PTK6 Regulates RHO GTPases, RAS GTPase and MAP kinases / Erythropoietin activates RAS / SHC-mediated cascade:FGFR1 / Signaling by FGFR4 in disease / striated muscle cell differentiation / Signaling by CSF3 (G-CSF) / FRS-mediated FGFR3 signaling / Signaling by FLT3 ITD and TKD mutants / FRS-mediated FGFR2 signaling / FRS-mediated FGFR4 signaling / p38MAPK events / FRS-mediated FGFR1 signaling / Signaling by FGFR3 in disease / Tie2 Signaling / protein-membrane adaptor activity / Signaling by FGFR2 in disease / Signaling by FLT3 fusion proteins / GRB2 events in EGFR signaling / SHC1 events in EGFR signaling / FLT3 Signaling / Signaling by FGFR1 in disease / EGFR Transactivation by Gastrin / NCAM signaling for neurite out-growth / CD209 (DC-SIGN) signaling / GRB2 events in ERBB2 signaling / Downstream signal transduction / response to glucocorticoid / Insulin receptor signalling cascade / SHC1 events in ERBB2 signaling / liver development / Constitutive Signaling by Overexpressed ERBB2 / Ras activation upon Ca2+ influx through NMDA receptor / Signaling by phosphorylated juxtamembrane, extracellular and kinase domain KIT mutants / VEGFR2 mediated cell proliferation / small monomeric GTPase / FCERI mediated MAPK activation / regulation of long-term neuronal synaptic plasticity / female pregnancy / Signaling by ERBB2 TMD/JMD mutants / Signaling by SCF-KIT / RAF activation / Constitutive Signaling by EGFRvIII / visual learning / Signaling by high-kinase activity BRAF mutants / Signaling by ERBB2 ECD mutants / MAP2K and MAPK activation / Signaling by ERBB2 KD Mutants / cytokine-mediated signaling pathway / cytoplasmic side of plasma membrane / gene expression / Signaling by RAF1 mutants / Signaling by CSF1 (M-CSF) in myeloid cells / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / Negative regulation of MAPK pathway / RAS processing / Regulation of RAS by GAPs / MAPK cascade / positive regulation of cellular senescence / Signaling by BRAF and RAF1 fusions / neuron apoptotic process / GDP binding Similarity search - Function
Small GTPase, Ras-type / Small GTPase Ras domain profile. / Ran (Ras-related nuclear proteins) /TC4 subfamily of small GTPases / Rho (Ras homology) subfamily of Ras-like small GTPases / Ras subfamily of RAS small GTPases / Small GTPase / Ras family / Rab subfamily of small GTPases / Small GTP-binding protein domain / P-loop containing nucleotide triphosphate hydrolases ...Small GTPase, Ras-type / Small GTPase Ras domain profile. / Ran (Ras-related nuclear proteins) /TC4 subfamily of small GTPases / Rho (Ras homology) subfamily of Ras-like small GTPases / Ras subfamily of RAS small GTPases / Small GTPase / Ras family / Rab subfamily of small GTPases / Small GTP-binding protein domain / P-loop containing nucleotide triphosphate hydrolases / Rossmann fold / P-loop containing nucleoside triphosphate hydrolase / 3-Layer(aba) Sandwich / Alpha Beta Similarity search - Domain/homology
Type: DECTRIS EIGER X 4M / Detector: PIXEL / Date: Dec 13, 2021
Radiation
Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray
Radiation wavelength
Wavelength: 1.045 Å / Relative weight: 1
Reflection
Resolution: 1.534→57.674 Å / Num. obs: 21593 / % possible obs: 95 % / Redundancy: 20.49 % Details: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last ...Details: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last updated 2020-04-13: http://mmcif.wwpdb.org/dictionaries/mmcif_pdbx_v50.dic/Index/) and the actual quantities provided by MRFANA (https://github.com/githubgphl/MRFANA) from the autoPROC package (https://www.globalphasing.com/autoproc/). In general, the mmCIF categories here should provide items that are currently used in the PDB archive. If there are alternatives, the one recommended by the PDB developers has been selected. The distinction between *_all and *_obs quantities is not always clear: often only one version is actively used within the PDB archive (or is the one recommended by PDB developers). The intention of distinguishing between classes of reflections before and after some kind of observation criterion was applied, can in principle be useful - but such criteria change in various ways throughout the data processing steps (rejection of overloaded or too partial reflections, outlier/misfit rejections during scaling etc) and there is no retrospect computation of data scaling/merging statistics for the reflections used in the final refinement (where another observation criterion might have been applied). Typical data processing will usually only provide one version of statistics at various stages and these are given in the recommended item here, irrespective of the "_all" and "_obs" connotation, see e.g. the use of _reflns.pdbx_Rmerge_I_obs, _reflns.pdbx_Rrim_I_all and _reflns.pdbx_Rpim_I_all. Please note that all statistics related to "merged intensities" (or "merging") are based on inverse-variance weighting of the individual measurements making up a symmetry-unique reflection. This is standard for several decades now, even if some of the dictionary definitions seem to suggest that a simple "mean" or "average" intensity is being used instead. R-values are always given for all symmetry-equivalent reflections following Friedel's law, i.e. Bijvoet pairs are not treated separately (since we want to describe the overall mean intensity and not the mean I(+) and I(-) here). The Rrim metric is identical to the Rmeas R-value and only differs in name. _reflns.pdbx_number_measured_all is the number of measured intensities just before the final merging step (at which point no additional rejection takes place). _reflns.number_obs is the number of symmetry-unique observations, i.e. the result of merging those measurements via inverse-variance weighting. _reflns.pdbx_netI_over_sigmaI is based on the merged intensities (_reflns.number_obs) as expected. _reflns.pdbx_redundancy is synonymous with "multiplicity". The per-shell item _reflns_shell.number_measured_all corresponds to the overall value _reflns.pdbx_number_measured_all. The per-shell item _reflns_shell.number_unique_all corresponds to the overall value _reflns.number_obs. The per-shell item _reflns_shell.percent_possible_all corresponds to the overall value _reflns.percent_possible_obs. The per-shell item _reflns_shell.meanI_over_sigI_obs corresponds to the overall value given as _reflns.pdbx_netI_over_sigmaI. But be aware of the incorrect definition of the former in the current dictionary! CC1/2: 0.999 / CC1/2 anomalous: -0.329 / Rmerge(I) obs: 0.104 / Rpim(I) all: 0.0232 / Rrim(I) all: 0.1065 / AbsDiff over sigma anomalous: 0.743 / Baniso tensor eigenvalue 1: 30.8 Å2 / Baniso tensor eigenvalue 2: 30.8 Å2 / Baniso tensor eigenvalue 3: 17.8 Å2 / Baniso tensor eigenvector 1 ortho1: 1 / Baniso tensor eigenvector 1 ortho2: 0 / Baniso tensor eigenvector 1 ortho3: 0 / Baniso tensor eigenvector 2 ortho1: 0 / Baniso tensor eigenvector 2 ortho2: 1 / Baniso tensor eigenvector 2 ortho3: 0 / Baniso tensor eigenvector 3 ortho1: 0 / Baniso tensor eigenvector 3 ortho2: 0 / Baniso tensor eigenvector 3 ortho3: 1 / Aniso diffraction limit 1: 1.856 Å / Aniso diffraction limit 2: 1.856 Å / Aniso diffraction limit 3: 1.499 Å / Aniso diffraction limit axis 1 ortho1: 1 / Aniso diffraction limit axis 1 ortho2: 0 / Aniso diffraction limit axis 1 ortho3: 0 / Aniso diffraction limit axis 2 ortho1: 0 / Aniso diffraction limit axis 2 ortho2: 1 / Aniso diffraction limit axis 2 ortho3: 0 / Aniso diffraction limit axis 3 ortho1: 0 / Aniso diffraction limit axis 3 ortho2: 0 / Aniso diffraction limit axis 3 ortho3: 1 / Net I/σ(I): 19.1 / Num. measured all: 442425 / Orthogonalization convention: pdb / % possible anomalous: 94.4 / % possible ellipsoidal: 95 / % possible ellipsoidal anomalous: 94.4 / % possible spherical: 66.2 / % possible spherical anomalous: 65.8 / Redundancy anomalous: 10.42
Reflection shell
Resolution (Å)
Redundancy (%)
Rmerge(I) obs
Mean I/σ(I) obs
Num. measured all
Num. measured obs
Num. unique all
Num. unique obs
CC1/2
CC1/2 anomalous
Rpim(I) all
Rrim(I) all
AbsDiff over sigma anomalous
% possible anomalous
% possible ellipsoidal
% possible ellipsoidal anomalous
% possible spherical
% possible spherical anomalous
Redundancy anomalous
% possible all
4.808-57.674
20.95
0.0358
71.52
22624
22624
1080
1080
0.999
-0.567
0.008
0.0367
0.818
99.5
99.9
99.5
99.9
99.5
10.98
99.9
3.819-4.808
21.99
0.0395
68.84
23796
23796
1082
1082
0.999
-0.449
0.0086
0.0404
0.75
100
100
100
100
100
11.28
100
3.326-3.819
22.23
0.0531
55.19
23893
23893
1075
1075
0.999
-0.354
0.0115
0.0543
0.784
100
100
100
100
100
11.32
100
3.02-3.326
22.38
0.0753
39.69
24123
24123
1078
1078
0.999
-0.307
0.0162
0.077
0.773
100
100
100
100
100
11.42
100
2.801-3.02
22.3
0.1073
29.98
24175
24175
1084
1084
0.997
-0.209
0.0233
0.1099
0.786
100
100
100
100
100
11.32
100
2.638-2.801
19.34
0.1406
20.45
20873
20873
1079
1079
0.996
-0.113
0.0327
0.1444
0.781
100
100
100
100
100
9.88
100
2.504-2.638
17.85
0.1696
16.62
19301
19301
1081
1081
0.994
-0.115
0.0412
0.1746
0.776
100
100
100
100
100
9.1
100
2.393-2.504
19.14
0.2144
14.31
20618
20618
1077
1077
0.993
-0.015
0.0503
0.2203
0.777
100
100
100
100
100
9.71
100
2.301-2.393
20.21
0.2533
12.5
21863
21863
1082
1082
0.994
-0.041
0.0577
0.2598
0.747
100
100
100
100
100
10.22
100
2.22-2.301
20.74
0.3371
10.17
22355
22355
1078
1078
0.987
-0.055
0.0757
0.3456
0.764
100
100
100
100
100
10.48
100
2.151-2.22
21.03
0.3993
8.71
22776
22776
1083
1083
0.986
-0.083
0.0889
0.4091
0.71
100
100
100
100
100
10.63
100
2.088-2.151
21.19
0.4913
7.22
22839
22839
1078
1078
0.979
0.021
0.1088
0.5033
0.754
100
100
100
100
100
10.71
100
2.035-2.088
21.27
0.6397
5.62
22908
22908
1077
1077
0.974
0.008
0.1411
0.6552
0.746
99.9
100
99.9
100
99.9
10.76
100
1.984-2.035
21.48
0.8072
4.51
23238
23238
1082
1082
0.95
-0.036
0.1771
0.8265
0.71
100
100
100
100
100
10.86
100
1.94-1.984
21.52
0.9657
3.81
23201
23201
1078
1078
0.94
-0.063
0.2115
0.9887
0.714
100
100
100
100
100
10.88
100
1.897-1.94
21.65
1.2256
3.03
23359
23359
1079
1079
0.902
-0.078
0.2673
1.2546
0.683
99.8
99.7
99.8
99.7
99.8
10.92
99.7
1.855-1.897
21.57
1.3791
2.69
23358
23358
1083
1083
0.895
-0.004
0.3008
1.4117
0.692
91
90.2
91
90.2
91
10.85
90.2
1.787-1.855
20.27
1.1842
3.05
21909
21909
1081
1081
0.894
-0.085
0.2663
1.214
0.724
72.3
72.4
72.3
49.3
50.3
10.15
72.4
1.694-1.787
18.88
1.3448
2.48
20255
20255
1073
1073
0.829
-0.059
0.3125
1.3811
0.679
73.7
73.9
73.7
30
30.5
9.46
73.9
1.534-1.694
13.81
1.5359
1.74
14961
14961
1083
1083
0.671
-0.024
0.4151
1.5931
0.704
73.8
81.9
73.8
12.9
11.9
7.41
81.9
-
Processing
Software
Name
Version
Classification
autoPROC
1.1.7
dataprocessing
XDS
Jan31, 2020
datareduction
Aimless
0.7.7
datascaling
pointless
1.12.12
datascaling
STARANISO
2.3.63
datascaling
BUSTER
2.11.8
refinement
PHASER
phasing
Refinement
Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 1.534→57.67 Å / Cor.coef. Fo:Fc: 0.941 / Cor.coef. Fo:Fc free: 0.932 / SU R Cruickshank DPI: 0.126 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.128 / SU Rfree Blow DPI: 0.116 / SU Rfree Cruickshank DPI: 0.116
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads PDBj
PDBj




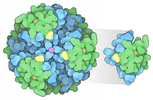







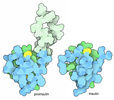




 Assembly
Assembly

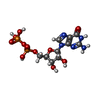


 Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM
Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION
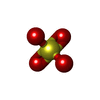
Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 96.063 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: SO4 / Feature type: SUBJECT OF INVESTIGATION
Mass: 96.063 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: SO4 / Feature type: SUBJECT OF INVESTIGATION Sample preparation
Sample preparation / Beamline: BL-1A / Wavelength: 1.045 Å
/ Beamline: BL-1A / Wavelength: 1.045 Å Processing
Processing