National Natural Science Foundation of China (NSFC)
31870740, 32171227, 31570738
中国
引用
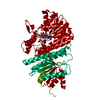
ジャーナル: mBio / 年: 2023 タイトル: Structural basis of a bi-functional malonyl-CoA reductase (MCR) from the photosynthetic green non-sulfur bacterium . 著者: Xin Zhang / Jiyu Xin / Zhiguo Wang / Wenping Wu / Yutong Liu / Zhenzhen Min / Yueyong Xin / Bing Liu / Jun He / Xingwei Zhang / Xiaoling Xu / 要旨: Malonyl-CoA reductase (MCR) is a NADPH-dependent bi-functional enzyme that performs alcohol dehydrogenase and aldehyde dehydrogenase (CoA-acylating) activities in the N- and C-terminal fragments, ...Malonyl-CoA reductase (MCR) is a NADPH-dependent bi-functional enzyme that performs alcohol dehydrogenase and aldehyde dehydrogenase (CoA-acylating) activities in the N- and C-terminal fragments, respectively. It catalyzes the two-step reduction of malonyl-CoA to 3-hydroxypropionate (3-HP), a key reaction in the autotrophic CO fixation cycles of green non-sulfur bacteria and the archaea . However, the structural basis underlying substrate selection, coordination, and the subsequent catalytic reactions of full-length MCR is largely unknown. For the first time, we here determined the structure of full-length MCR from the photosynthetic green non-sulfur bacterium (MCR) at 3.35 Å resolution. Furthermore, we determined the crystal structures of the N- and C-terminal fragments bound with reaction intermediates NADP and malonate semialdehyde (MSA) at 2.0 Å and 2.3 Å, respectively, and elucidated the catalytic mechanisms using a combination of molecular dynamics simulations and enzymatic analyses. Full-length MCR was a homodimer of two cross-interlocked subunits, each containing four tandemly arranged short-chain dehydrogenase/reductase (SDR) domains. Only the catalytic domains SDR1 and SDR3 incorporated additional secondary structures that changed with NADP-MSA binding. The substrate, malonyl-CoA, was immobilized in the substrate-binding pocket of SDR3 through coordination with Arg1164 and Arg799 of SDR4 and the extra domain, respectively. Malonyl-CoA was successively reduced through protonation by the Tyr743-Arg746 pair in SDR3 and the catalytic triad (Thr165-Tyr178-Lys182) in SDR1 after nucleophilic attack from NADPH hydrides. IMPORTANCE The bi-functional MCR catalyzes NADPH-dependent reduction of malonyl-CoA to 3-HP, an important metabolic intermediate and platform chemical, from biomass. The individual MCR-N and MCR-C fragments, which contain the alcohol dehydrogenase and aldehyde dehydrogenase (CoA-acylating) activities, respectively, have previously been structurally investigated and reconstructed into a malonyl-CoA pathway for the biosynthetic production of 3-HP. However, no structural information for full-length MCR has been available to illustrate the catalytic mechanism of this enzyme, which greatly limits our capacity to increase the 3-HP yield of recombinant strains. Here, we report the cryo-electron microscopy structure of full-length MCR for the first time and elucidate the mechanisms underlying substrate selection, coordination, and catalysis in the bi-functional MCR. These findings provide a structural and mechanistic basis for enzyme engineering and biosynthetic applications of the 3-HP carbon fixation pathways.
分子量: 73024.969 Da / 分子数: 1 / 断片: C terminal domain / 由来タイプ: 組換発現 詳細: The amino acid residues Met563-Ser566 are sequences derived from the expression vector, and His567-His572 are expression tag for purification of the target protein 由来: (組換発現) Roseiflexus castenholzii DSM 13941 (バクテリア) 株: DSM 13941 / HLO8 / 遺伝子: Rcas_2929 / 発現宿主: Escherichia coli BL21(DE3) (大腸菌) / 参照: UniProt: A7NN59
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Roseiflexus castenholzii DSM 13941 (バクテリア)
Roseiflexus castenholzii DSM 13941 (バクテリア) X線回折 /
X線回折 /  データ登録者
データ登録者 中国, 1件
中国, 1件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード

 PDBj
PDBj
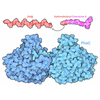
 集合体
集合体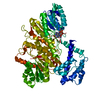
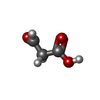
 分子量: 88.062 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H4O3 / タイプ: SUBJECT OF INVESTIGATION
分子量: 88.062 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H4O3 / タイプ: SUBJECT OF INVESTIGATION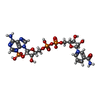
 分子量: 743.405 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C21H28N7O17P3 / タイプ: SUBJECT OF INVESTIGATION
分子量: 743.405 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C21H28N7O17P3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 18.015 Da / 分子数: 81 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 81 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析