 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8.0E+15 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | A computationally stabilized hMPV F protein | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | VIRAL PROTEIN / Human metapneumovirus / fusion protein / computational stabilization | |||||||||
| Function / homology |  Function and homology information Function and homology informationfusion of virus membrane with host plasma membrane / viral envelope / symbiont entry into host cell / host cell plasma membrane / virion membrane Similarity search - Function | |||||||||
| Biological species |  Human metapneumovirusEnterobacteria phage T2 (virus) Human metapneumovirusEnterobacteria phage T2 (virus) | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.41 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.41 Å | |||||||||
 Authors Authors | Huang, J. / Gonzalez, K. / Mousa, J. / Strauch, E. | |||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: A general computational design strategy for stabilizing viral class I fusion proteins. Authors: Karen J Gonzalez / Jiachen Huang / Miria F Criado / Avik Banerjee / Stephen M Tompkins / Jarrod J Mousa / Eva-Maria Strauch / Abstract: Many pathogenic viruses rely on class I fusion proteins to fuse their viral membrane with the host cell membrane. To drive the fusion process, class I fusion proteins undergo an irreversible ...Many pathogenic viruses rely on class I fusion proteins to fuse their viral membrane with the host cell membrane. To drive the fusion process, class I fusion proteins undergo an irreversible conformational change from a metastable prefusion state to an energetically more stable postfusion state. Mounting evidence underscores that antibodies targeting the prefusion conformation are the most potent, making it a compelling vaccine candidate. Here, we establish a computational design protocol that stabilizes the prefusion state while destabilizing the postfusion conformation. With this protocol, we stabilize the fusion proteins of the RSV, hMPV, and SARS-CoV-2 viruses, testing fewer than a handful of designs. The solved structures of these designed proteins from all three viruses evidence the atomic accuracy of our approach. Furthermore, the humoral response of the redesigned RSV F protein compares to that of the recently approved vaccine in a mouse model. While the parallel design of two conformations allows the identification of energetically sub-optimal positions for one conformation, our protocol also reveals diverse molecular strategies for stabilization. Given the clinical significance of viruses using class I fusion proteins, our algorithm can substantially contribute to vaccine development by reducing the time and resources needed to optimize these immunogens. #1: Journal: bioRxiv / Year: 2023 Title: A general computational design strategy for stabilizing viral class I fusion proteins. Authors: Karen J Gonzalez / Jiachen Huang / Miria F Criado / Avik Banerjee / Stephen Tompkins / Jarrod J Mousa / Eva-Maria Strauch / Abstract: Many pathogenic viruses, including influenza virus, Ebola virus, coronaviruses, and Pneumoviruses, rely on class I fusion proteins to fuse viral and cellular membranes. To drive the fusion process, ...Many pathogenic viruses, including influenza virus, Ebola virus, coronaviruses, and Pneumoviruses, rely on class I fusion proteins to fuse viral and cellular membranes. To drive the fusion process, class I fusion proteins undergo an irreversible conformational change from a metastable prefusion state to an energetically more favorable and stable postfusion state. An increasing amount of evidence exists highlighting that antibodies targeting the prefusion conformation are the most potent. However, many mutations have to be evaluated before identifying prefusion-stabilizing substitutions. We therefore established a computational design protocol that stabilizes the prefusion state while destabilizing the postfusion conformation. As a proof of concept, we applied this principle to the fusion protein of the RSV, hMPV, and SARS-CoV-2 viruses. For each protein, we tested less than a handful of designs to identify stable versions. Solved structures of designed proteins from the three different viruses evidenced the atomic accuracy of our approach. Furthermore, the immunological response of the RSV F design compared to a current clinical candidate in a mouse model. While the parallel design of two conformations allows identifying and selectively modifying energetically less optimized positions for one conformation, our protocol also reveals diverse molecular strategies for stabilization. We recaptured many approaches previously introduced manually for the stabilization of viral surface proteins, such as cavity-filling, optimization of polar interactions, as well as postfusion-disruptive strategies. Using our approach, it is possible to focus on the most impacting mutations and potentially preserve the immunogen as closely as possible to its native version. The latter is important as sequence re-design can cause perturbations to B and T cell epitopes. Given the clinical significance of viruses using class I fusion proteins, our algorithm can substantially contribute to vaccine development by reducing the time and resources needed to optimize these immunogens. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8e15.cif.gz | 105.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8e15.ent.gz | 75.9 KB | Display | PDB format |
| PDBx/mmJSON format | 8e15.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/e1/8e15ftp://data.pdbj.org/pub/pdb/validation_reports/e1/8e15 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  7tn1C 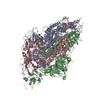 8fezC  5wb0S S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj

- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 11687.245 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Human metapneumovirus / Cell line (production host): 293 / Production host:  Homo sapiens (human) / References: UniProt: Q8B9P0 Homo sapiens (human) / References: UniProt: Q8B9P0 | ||||||
|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 45999.160 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Human metapneumovirus, (gene. exp.) Enterobacteria phage T2 (virus)Gene: F, wac, EcT2_00172 / Cell line (production host): 293 / Production host: Homo sapiens (human) / References: UniProt: Q8B9P0, UniProt: Q76VI8 | ||||||
| #3: Polysaccharide | beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta- ...beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose Source method: isolated from a genetically manipulated source | ||||||
| #4: Sugar | 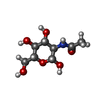  Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C8H15NO6 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C8H15NO6#5: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 11 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 11 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | Has protein modification | Y | |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 4.09 Å3/Da / Density % sol: 69.91 % |
|---|---|
| Crystal grow | Temperature: 298 K / Method: vapor diffusion, sitting drop Details: 0.1 M Sodium acetate trihydrate pH 4.6, 2.0 M Sodium formate |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS / Beamline: 21-ID-D / Wavelength: 1 Å |
| Detector | Type: DECTRIS EIGER X 16M / Detector: PIXEL / Date: Apr 21, 2022 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 2.41→47.62 Å / Num. obs: 36363 / % possible obs: 99.91 % / Redundancy: 2 % / Biso Wilson estimate: 66.92 Å2 / CC1/2: 0.999 / CC star: 1 / Net I/σ(I): 9.18 |
| Reflection shell | Resolution: 2.41→2.496 Å / Redundancy: 2 % / Mean I/σ(I) obs: 0.78 / Num. unique obs: 3606 / CC1/2: 0.593 / CC star: 0.863 / % possible all: 99.86 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 5wb0 Resolution: 2.41→47.62 Å / SU ML: 0.3364 / Cross valid method: FREE R-VALUE / σ(F): 1.34 / Phase error: 27.647 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.1 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 70.9 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.41→47.62 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
