positive regulation of I-kappaB phosphorylation / UFM1 ligase activity / UFM1-modified protein reader activity / positive regulation of reticulophagy / UFM1 conjugating enzyme activity / UFM1 transferase activity / positive regulation of protein localization to endoplasmic reticulum / positive regulation of proteolysis involved in protein catabolic process / protein K69-linked ufmylation / protein ufmylation ...positive regulation of I-kappaB phosphorylation / UFM1 ligase activity / UFM1-modified protein reader activity / positive regulation of reticulophagy / UFM1 conjugating enzyme activity / UFM1 transferase activity / positive regulation of protein localization to endoplasmic reticulum / positive regulation of proteolysis involved in protein catabolic process / protein K69-linked ufmylation / protein ufmylation / positive regulation of plasma cell differentiation / negative regulation of IRE1-mediated unfolded protein response / regulation of proteasomal ubiquitin-dependent protein catabolic process / regulation of type II interferon production / positive regulation of cell cycle G1/S phase transition / negative regulation of T cell mediated immune response to tumor cell / protein localization to endoplasmic reticulum / negative regulation of T cell activation / regulation of intracellular estrogen receptor signaling pathway / positive regulation of proteasomal protein catabolic process / cartilage development / 転移酵素; アシル基を移すもの; アミノアシル基を移すもの / ribosome disassembly / response to L-glutamate / regulation of canonical NF-kappaB signal transduction / reticulophagy / negative regulation of NF-kappaB transcription factor activity / negative regulation of PERK-mediated unfolded protein response / ubiquitin-like protein ligase binding / positive regulation of glial cell proliferation / RHOA GTPase cycle / hematopoietic stem cell differentiation / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / negative regulation of protein ubiquitination / positive regulation of autophagy / rescue of stalled ribosome / response to endoplasmic reticulum stress / DNA damage checkpoint signaling / erythrocyte differentiation / positive regulation of NF-kappaB transcription factor activity / brain development / regulation of protein stability / osteoblast differentiation / Antigen processing: Ubiquitination & Proteasome degradation / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / regulation of protein localization / site of double-strand break / regulation of inflammatory response / mitochondrial outer membrane / positive regulation of canonical NF-kappaB signal transduction / neuron projection / protein stabilization / positive regulation of cell migration / negative regulation of gene expression / DNA repair / positive regulation of cell population proliferation / DNA damage response / positive regulation of gene expression / endoplasmic reticulum membrane / protein kinase binding / negative regulation of apoptotic process / nucleolus / endoplasmic reticulum / positive regulation of transcription by RNA polymerase II / protein-containing complex / extracellular exosome / nucleus / membrane / cytoplasm / cytosol 類似検索 - 分子機能
Biotechnology and Biological Sciences Research Council (BBSRC)
BB/W007401/1
英国
引用
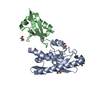
ジャーナル: Nature / 年: 2024 タイトル: The UFM1 E3 ligase recognizes and releases 60S ribosomes from ER translocons. 著者: Linda Makhlouf / Joshua J Peter / Helge M Magnussen / Rohan Thakur / David Millrine / Thomas C Minshull / Grace Harrison / Joby Varghese / Frederic Lamoliatte / Martina Foglizzo / Thomas ...著者: Linda Makhlouf / Joshua J Peter / Helge M Magnussen / Rohan Thakur / David Millrine / Thomas C Minshull / Grace Harrison / Joby Varghese / Frederic Lamoliatte / Martina Foglizzo / Thomas Macartney / Antonio N Calabrese / Elton Zeqiraj / Yogesh Kulathu / 要旨: Stalled ribosomes at the endoplasmic reticulum (ER) are covalently modified with the ubiquitin-like protein UFM1 on the 60S ribosomal subunit protein RPL26 (also known as uL24). This modification, ...Stalled ribosomes at the endoplasmic reticulum (ER) are covalently modified with the ubiquitin-like protein UFM1 on the 60S ribosomal subunit protein RPL26 (also known as uL24). This modification, which is known as UFMylation, is orchestrated by the UFM1 ribosome E3 ligase (UREL) complex, comprising UFL1, UFBP1 and CDK5RAP3 (ref. ). However, the catalytic mechanism of UREL and the functional consequences of UFMylation are unclear. Here we present cryo-electron microscopy structures of UREL bound to 60S ribosomes, revealing the basis of its substrate specificity. UREL wraps around the 60S subunit to form a C-shaped clamp architecture that blocks the tRNA-binding sites at one end, and the peptide exit tunnel at the other. A UFL1 loop inserts into and remodels the peptidyl transferase centre. These features of UREL suggest a crucial function for UFMylation in the release and recycling of stalled or terminated ribosomes from the ER membrane. In the absence of functional UREL, 60S-SEC61 translocon complexes accumulate at the ER membrane, demonstrating that UFMylation is necessary for releasing SEC61 from 60S subunits. Notably, this release is facilitated by a functional switch of UREL from a 'writer' to a 'reader' module that recognizes its product-UFMylated 60S ribosomes. Collectively, we identify a fundamental role for UREL in dissociating 60S subunits from the SEC61 translocon and the basis for UFMylation in regulating protein homeostasis at the ER.
E3UFM1-proteinligase1 / E3 UFM1-protein transferase 1 / Multiple alpha-helix protein located at ER / Novel LZAP-binding ...E3 UFM1-protein transferase 1 / Multiple alpha-helix protein located at ER / Novel LZAP-binding protein / Regulator of C53/LZAP and DDRGK1
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 英国, 1件
英国, 1件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード


 PDBj
PDBj


 集合体
集合体



 分子量: 18.015 Da / 分子数: 7 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 7 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 / ビームライン: ID23-2 / 波長: 0.873128 Å
/ ビームライン: ID23-2 / 波長: 0.873128 Å 解析
解析