ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7unc | ||||||
|---|---|---|---|---|---|---|---|
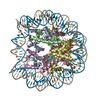
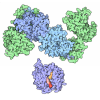
| タイトル | Pol II-DSIF-SPT6-PAF1c-TFIIS complex with rewrapped nucleosome | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | TRANSCRIPTION / Chromatin / nucleosome / intermediate | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報blastocyst growth / positive regulation of mRNA 3'-end processing / Ski complex / RNA polymerase II C-terminal domain phosphoserine binding / mRNA decay by 3' to 5' exoribonuclease / Cdc73/Paf1 complex / regulation of isotype switching / negative regulation of DNA-templated transcription, elongation / inner cell mass cell differentiation / nuclear-transcribed mRNA catabolic process, 3'-5' exonucleolytic nonsense-mediated decay ...blastocyst growth / positive regulation of mRNA 3'-end processing / Ski complex / RNA polymerase II C-terminal domain phosphoserine binding / mRNA decay by 3' to 5' exoribonuclease / Cdc73/Paf1 complex / regulation of isotype switching / negative regulation of DNA-templated transcription, elongation / inner cell mass cell differentiation / nuclear-transcribed mRNA catabolic process, 3'-5' exonucleolytic nonsense-mediated decay / regulation of muscle cell differentiation / regulation of mRNA export from nucleus / endodermal cell fate commitment / negative regulation of myeloid cell differentiation / DSIF complex / positive regulation of cell cycle G1/S phase transition / trophectodermal cell differentiation / blastocyst hatching / regulation of transcription elongation by RNA polymerase II / regulation of mRNA processing / nucleosome organization / blastocyst formation / nuclear lumen / mRNA 3'-end processing / positive regulation of DNA-templated transcription, elongation / Abortive elongation of HIV-1 transcript in the absence of Tat / transcription elongation-coupled chromatin remodeling / stem cell population maintenance / transcription factor TFIID complex / interleukin-6-mediated signaling pathway / negative regulation of G1/S transition of mitotic cell cycle / RNA Pol II CTD phosphorylation and interaction with CE during HIV infection / RNA Pol II CTD phosphorylation and interaction with CE / negative regulation of gene expression, epigenetic / Formation of the Early Elongation Complex / Formation of the HIV-1 Early Elongation Complex / mRNA Capping / RNA polymerase II complex binding / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / positive regulation of macroautophagy / RNA polymerase II transcribes snRNA genes / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / positive regulation of Wnt signaling pathway / cell surface receptor signaling pathway via JAK-STAT / protein localization to nucleus / mRNA transport / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / transcription by RNA polymerase III / transcription by RNA polymerase I / RNA polymerase I complex / Formation of HIV elongation complex in the absence of HIV Tat / RNA polymerase III complex / RNA polymerase II, core complex / tRNA transcription by RNA polymerase III / nucleosome binding / : / RNA Polymerase II Transcription Elongation / negative regulation of fibroblast proliferation / Formation of RNA Pol II elongation complex / RNA Polymerase II Pre-transcription Events / rescue of stalled ribosome / DNA-directed RNA polymerase complex / DNA-directed RNA polymerase activity / SH2 domain binding / RNA splicing / transcription elongation factor complex / positive regulation of transcription elongation by RNA polymerase II / TP53 Regulates Transcription of DNA Repair Genes / DNA-templated transcription initiation / regulation of cell growth / transcription elongation by RNA polymerase II / Formation of the beta-catenin:TCF transactivating complex / Hedgehog 'on' state / euchromatin / protein destabilization / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / ribonucleoside binding / Formation of TC-NER Pre-Incision Complex / Wnt signaling pathway / : / : / : / : / : / : / fibrillar center / DNA-directed RNA polymerase / mRNA processing / Dual incision in TC-NER / negative regulation of epithelial cell proliferation / Gap-filling DNA repair synthesis and ligation in TC-NER / structural constituent of chromatin / nucleosome / heterochromatin formation / E3 ubiquitin ligases ubiquitinate target proteins / nucleosome assembly / single-stranded DNA binding 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) synthetic construct (人工物) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3 Å | ||||||
 データ登録者 データ登録者 | Filipovski, M. / Vos, S.M. / Farnung, L. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Science / 年: 2022 タイトル: Structural basis of nucleosome retention during transcription elongation. 著者: Martin Filipovski / Jelly H M Soffers / Seychelle M Vos / Lucas Farnung / 要旨: In eukaryotes, RNA polymerase (Pol) II transcribes chromatin and must move past nucleosomes, often resulting in nucleosome displacement. How Pol II unwraps the DNA from nucleosomes to allow ...In eukaryotes, RNA polymerase (Pol) II transcribes chromatin and must move past nucleosomes, often resulting in nucleosome displacement. How Pol II unwraps the DNA from nucleosomes to allow transcription and how DNA rewraps to retain nucleosomes has been unclear. Here, we report the 3.0-angstrom cryo-electron microscopy structure of a mammalian Pol II-DSIF-SPT6-PAF1c-TFIIS-nucleosome complex stalled 54 base pairs within the nucleosome. The structure provides a mechanistic basis for nucleosome retention during transcription elongation where upstream DNA emerging from the Pol II cleft has rewrapped the proximal side of the nucleosome. The structure uncovers a direct role for Pol II and transcription elongation factors in nucleosome retention and explains how nucleosomes are retained to prevent the disruption of chromatin structure across actively transcribed genes. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7unc.cif.gz | 2.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7unc.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7unc.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7unc_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7unc_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 7unc_validation.xml.gz | 177.5 KB | 表示 | |
| CIF形式データ | 7unc_validation.cif.gz | 301.9 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/un/7uncftp://data.pdbj.org/pub/pdb/validation_reports/un/7unc | HTTPS FTP |
-関連構造データ


-リンク
 PDBj
PDBj





































































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA-directed RNA polymerase ... , 5種, 5分子 ABCEG
| #1: タンパク質 | 分子量: 219049.500 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 142426.125 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質 | 分子量: 31439.074 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 24644.318 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 19314.283 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 12種, 16分子 DFHIJKWXaebfcgdh
| #4: タンパク質 | 分子量: 20962.621 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
|---|---|---|---|---|---|---|---|
| #6: タンパク質 | 分子量: 14477.001 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #8: タンパク質 | 分子量: 17162.273 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #9: タンパク質 | 分子量: 14541.221 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #10: タンパク質 | 分子量: 7655.123 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #11: タンパク質 | 分子量: 13310.284 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #22: タンパク質 | 分子量: 33617.465 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q9GZS3 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q9GZS3 | ||||||
| #23: タンパク質 | 分子量: 60673.539 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CDC73, C1orf28, HRPT2 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q6P1J9 | ||||||
| #25: タンパク質 | 分子量: 15435.126 Da / 分子数: 2 / Mutation: G102A / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主:  #26: タンパク質 | 分子量: 11394.426 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: #27: タンパク質 | 分子量: 14109.436 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: hist1h2aj, LOC494591, XELAEV_18003602mg / 発現宿主: #28: タンパク質 | 分子量: 13655.948 Da / 分子数: 2 / Mutation: S29T / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-RNA polymerase ... , 2種, 2分子 LV
| #12: タンパク質 | 分子量: 7018.244 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #21: タンパク質 | 分子量: 60052.672 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PAF1, PD2 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q8N7H5 |
-Transcription elongation factor ... , 3種, 3分子 MOZ
| #13: タンパク質 | 分子量: 199602.969 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUPT6H, KIAA0162, SPT6H / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q7KZ85 |
|---|---|
| #15: タンパク質 | 分子量: 34294.980 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: TCEA1, GTF2S, TFIIS / 発現宿主: |
| #24: タンパク質 | 分子量: 121225.477 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUPT5H, SPT5, SPT5H / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: O00267 |
-DNA鎖 , 2種, 2分子 NT
| #14: DNA鎖 | 分子量: 64763.039 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #19: DNA鎖 | 分子量: 66186.398 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-RNA鎖 , 1種, 1分子 P
| #16: RNA鎖 | 分子量: 5004.936 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|
-RNA polymerase-associated protein ... , 3種, 3分子 QRU
| #17: タンパク質 | 分子量: 134510.203 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CTR9, KIAA0155, SH2BP1 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q6PD62 |
|---|---|
| #18: タンパク質 | 分子量: 80733.016 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RTF1, KIAA0252 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q92541 |
| #20: タンパク質 | 分子量: 75514.172 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LEO1, RDL / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q8WVC0 |
-非ポリマー , 2種, 9分子 

| #29: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#30: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Pol II-DSIF-SPT6-PAF1c-TFIIS complex with rewrapped nucleosome タイプ: COMPLEX / Entity ID: #1-#28 / 由来: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | 詳細: 15 mA with 10 s hold time / グリッドの材料: COPPER / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/1 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 2000 nm / 最小 デフォーカス(公称値): 500 nm |
| 撮影 | 電子線照射量: 52 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
| 電子光学装置 | エネルギーフィルタースリット幅: 20 eV / 位相板: VOLTA PHASE PLATE |
- 解析
解析
| ソフトウェア | 名称: UCSF ChimeraX / バージョン: 1.4/v9 / 分類: モデル構築 / URL: https://www.rbvi.ucsf.edu/chimerax/ / Os: macOS / タイプ: package | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 105420 / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 189.36 / プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||
| 原子モデル構築 |
|