 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7ra7 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of rabbit anti-HIV Fab 11A | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | IMMUNE SYSTEM / antibody antigen neutralizing antibody HIV | |||||||||
| Function / homology | Immunoglobulins / Immunoglobulin-like / Sandwich / Mainly Beta Function and homology information Function and homology information | |||||||||
| Biological species |  | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.2 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.2 Å | |||||||||
 Authors Authors | Oyen, D. / Wilson, I.A. | |||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||
 Citation Citation | Journal: To Be Published Title: Limited breadth of anti-HIV Env glycan hole antibodies is further hindered by strain-specific peptide interactions Authors: Ozorowski, G. / Cottrell, C.A. / Oyen, D. / de Val, N. / Copps, J. / Scaring, N. / Polveroni, T.M. / Wilson, I.A. / Ward, A.B. #1: Journal: Immunity / Year: 2018Title: Electron-Microscopy-Based Epitope Mapping Defines Specificities of Polyclonal Antibodies Elicited during HIV-1 BG505 Envelope Trimer Immunization. Authors: Matteo Bianchi / Hannah L Turner / Bartek Nogal / Christopher A Cottrell / David Oyen / Matthias Pauthner / Raiza Bastidas / Rebecca Nedellec / Laura E McCoy / Ian A Wilson / Dennis R Burton ...Authors: Matteo Bianchi / Hannah L Turner / Bartek Nogal / Christopher A Cottrell / David Oyen / Matthias Pauthner / Raiza Bastidas / Rebecca Nedellec / Laura E McCoy / Ian A Wilson / Dennis R Burton / Andrew B Ward / Lars Hangartner /  Abstract: Characterizing polyclonal antibody responses via currently available methods is inherently complex and difficult. Mapping epitopes in an immune response is typically incomplete, which creates a ...Characterizing polyclonal antibody responses via currently available methods is inherently complex and difficult. Mapping epitopes in an immune response is typically incomplete, which creates a barrier to fully understanding the humoral response to antigens and hinders rational vaccine design efforts. Here, we describe a method of characterizing polyclonal responses by using electron microscopy, and we applied this method to the immunization of rabbits with an HIV-1 envelope glycoprotein vaccine candidate, BG505 SOSIP.664. We detected known epitopes within the polyclonal sera and revealed how antibody responses evolved during the prime-boosting strategy to ultimately result in a neutralizing antibody response. We uncovered previously unidentified epitopes, including an epitope proximal to one recognized by human broadly neutralizing antibodies as well as potentially distracting non-neutralizing epitopes. Our method provides an efficient and semiquantitative map of epitopes that are targeted in a polyclonal antibody response and should be of widespread utility in vaccine and infection studies. #2: Journal: Cell Rep / Year: 2016Title: Holes in the Glycan Shield of the Native HIV Envelope Are a Target of Trimer-Elicited Neutralizing Antibodies. Authors: Laura E McCoy / Marit J van Gils / Gabriel Ozorowski / Terrence Messmer / Bryan Briney / James E Voss / Daniel W Kulp / Matthew S Macauley / Devin Sok / Matthias Pauthner / Sergey Menis / ...Authors: Laura E McCoy / Marit J van Gils / Gabriel Ozorowski / Terrence Messmer / Bryan Briney / James E Voss / Daniel W Kulp / Matthew S Macauley / Devin Sok / Matthias Pauthner / Sergey Menis / Christopher A Cottrell / Jonathan L Torres / Jessica Hsueh / William R Schief / Ian A Wilson / Andrew B Ward / Rogier W Sanders / Dennis R Burton /  Abstract: A major advance in the search for an HIV vaccine has been the development of a near-native Envelope trimer (BG505 SOSIP.664) that can induce robust autologous Tier 2 neutralization. Here, potently ...A major advance in the search for an HIV vaccine has been the development of a near-native Envelope trimer (BG505 SOSIP.664) that can induce robust autologous Tier 2 neutralization. Here, potently neutralizing monoclonal antibodies (nAbs) from rabbits immunized with BG505 SOSIP.664 are shown to recognize an immunodominant region of gp120 centered on residue 241. Residue 241 occupies a hole in the glycan defenses of the BG505 isolate, with fewer than 3% of global isolates lacking a glycan site at this position. However, at least one conserved glycan site is missing in 89% of viruses, suggesting the presence of glycan holes in most HIV isolates. Serum evidence is consistent with targeting of holes in natural infection. The immunogenic nature of breaches in the glycan shield has been under-appreciated in previous attempts to understand autologous neutralizing antibody responses and has important potential consequences for HIV vaccine design. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7ra7.cif.gz | 382.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7ra7.ent.gz | 251.8 KB | Display | PDB format |
| PDBx/mmJSON format | 7ra7.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ra/7ra7ftp://data.pdbj.org/pub/pdb/validation_reports/ra/7ra7 | HTTPS FTP |
|---|
-Related structure data





-Links
 PDBj
PDBj
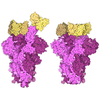
- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||||
| 2 | 
| ||||||||||||
| Unit cell |
|
-Components
| #1: Antibody | Mass: 24072.021 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  Homo sapiens (human) Homo sapiens (human)#2: Antibody | Mass: 22777.102 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)#3: Chemical | ChemComp-SO4 / 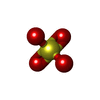  Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4 Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4#4: Chemical | ChemComp-1PE / | 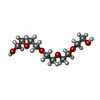  Mass: 238.278 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H22O6 / Comment: precipitant*YM Mass: 238.278 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H22O6 / Comment: precipitant*YM#5: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 374 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 374 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | Has protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.41 Å3/Da / Density % sol: 48.95 % |
|---|---|
| Crystal grow | Temperature: 298 K / Method: vapor diffusion, sitting drop / pH: 4.5 Details: 0.1M sodium acetate, 0.2M lithium sulfate, 30% PEG8000 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS / Beamline: 23-ID-D / Wavelength: 1.03315 Å |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Oct 15, 2016 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1.03315 Å / Relative weight: 1 |
| Reflection | Resolution: 2.2→45.9 Å / Num. obs: 44732 / % possible obs: 98.7 % / Observed criterion σ(F): 0 / Observed criterion σ(I): -3 / Redundancy: 6.4 % / Biso Wilson estimate: 38.17 Å2 / CC1/2: 0.86 / Rmerge(I) obs: 0.185 / Rpim(I) all: 0.078 / Rrim(I) all: 0.201 / Net I/σ(I): 9 |
| Reflection shell | Resolution: 2.2→2.24 Å / Redundancy: 5.2 % / Rmerge(I) obs: 1.2 / Mean I/σ(I) obs: 1 / Num. unique obs: 2195 / CC1/2: 0.56 / Rpim(I) all: 0.55 / Rrim(I) all: 1.3 / % possible all: 96.5 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: homology model Resolution: 2.2→45.9 Å / SU ML: 0.323 / Cross valid method: FREE R-VALUE / σ(F): 1.34 / Phase error: 29.2954 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 41.27 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.2→45.9 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
