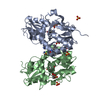
Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
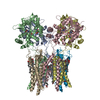
Yorodumi- PDB-7qhh: Desensitized state of GluA1/2 AMPA receptor in complex with TARP-... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7qhh | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Desensitized state of GluA1/2 AMPA receptor in complex with TARP-gamma 8 (TMD-LBD) | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | MEMBRANE PROTEIN / glutamate / AMPA receptor / TARPs | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationPhase 0 - rapid depolarisation / Phase 2 - plateau phase / Cargo concentration in the ER / cellular response to ammonium ion / axonal spine / positive regulation of locomotion involved in locomotory behavior / COPII-mediated vesicle transport / positive regulation of membrane potential / response to sucrose / L-type voltage-gated calcium channel complex ...Phase 0 - rapid depolarisation / Phase 2 - plateau phase / Cargo concentration in the ER / cellular response to ammonium ion / axonal spine / positive regulation of locomotion involved in locomotory behavior / COPII-mediated vesicle transport / positive regulation of membrane potential / response to sucrose / L-type voltage-gated calcium channel complex / regulation of monoatomic ion transmembrane transport / myosin V binding / positive regulation of AMPA receptor activity / LGI-ADAM interactions / neuron spine / Trafficking of AMPA receptors / postsynaptic neurotransmitter receptor diffusion trapping / proximal dendrite / protein phosphatase 2B binding / response to arsenic-containing substance / regulation of AMPA receptor activity / cellular response to L-glutamate / channel regulator activity / cellular response to dsRNA / long-term synaptic depression / ligand-gated calcium channel activity / dendritic spine membrane / beta-2 adrenergic receptor binding / Synaptic adhesion-like molecules / cellular response to peptide hormone stimulus / spine synapse / dendritic spine neck / dendritic spine cytoplasm / dendritic spine head / cellular response to amine stimulus / response to morphine / response to psychosocial stress / peptide hormone receptor binding / Activation of AMPA receptors / ligand-gated monoatomic cation channel activity / perisynaptic space / spinal cord development / neuronal cell body membrane / Trafficking of GluR2-containing AMPA receptors / protein kinase A binding / response to lithium ion / AMPA glutamate receptor activity / AMPA glutamate receptor clustering / behavioral response to pain / transmission of nerve impulse / regulation of receptor recycling / kainate selective glutamate receptor activity / immunoglobulin binding / adenylate cyclase binding / AMPA glutamate receptor complex / response to electrical stimulus / extracellularly glutamate-gated ion channel activity / cellular response to glycine / ionotropic glutamate receptor complex / asymmetric synapse / Unblocking of NMDA receptors, glutamate binding and activation / G-protein alpha-subunit binding / glutamate receptor binding / positive regulation of synaptic transmission / conditioned place preference / long-term memory / regulation of postsynaptic membrane neurotransmitter receptor levels / neuronal action potential / voltage-gated calcium channel activity / postsynaptic density, intracellular component / regulation of synaptic transmission, glutamatergic / response to fungicide / extracellular ligand-gated monoatomic ion channel activity / cytoskeletal protein binding / glutamate-gated receptor activity / cellular response to brain-derived neurotrophic factor stimulus / regulation of long-term synaptic depression / positive regulation of synaptic transmission, glutamatergic / glutamate-gated calcium ion channel activity / somatodendritic compartment / synapse assembly / presynaptic active zone membrane / ionotropic glutamate receptor signaling pathway / ionotropic glutamate receptor binding / dendrite cytoplasm / dendrite membrane / excitatory synapse / ligand-gated monoatomic ion channel activity involved in regulation of presynaptic membrane potential / positive regulation of excitatory postsynaptic potential / cellular response to amino acid stimulus / dendritic shaft / SNARE binding / synaptic membrane / response to cocaine / PDZ domain binding / calcium channel regulator activity / neuromuscular junction / establishment of protein localization / protein tetramerization / synaptic transmission, glutamatergic Similarity search - Function | ||||||||||||
| Biological species |  | ||||||||||||
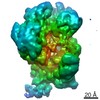
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | ||||||||||||
 Authors Authors | Herguedas, B. / Kohegyi, B. / Dohrke, J.N. / Watson, J.F. / Zhang, D. / Ho, H. / Shaikh, S. / Lape, R. / Krieger, J.M. / Greger, I.H. | ||||||||||||
| Funding support |  United Kingdom, 3items United Kingdom, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2022 Title: Mechanisms underlying TARP modulation of the GluA1/2-γ8 AMPA receptor. Authors: Beatriz Herguedas / Bianka K Kohegyi / Jan-Niklas Dohrke / Jake F Watson / Danyang Zhang / Hinze Ho / Saher A Shaikh / Remigijus Lape / James M Krieger / Ingo H Greger /    Abstract: AMPA-type glutamate receptors (AMPARs) mediate rapid signal transmission at excitatory synapses in the brain. Glutamate binding to the receptor's ligand-binding domains (LBDs) leads to ion channel ...AMPA-type glutamate receptors (AMPARs) mediate rapid signal transmission at excitatory synapses in the brain. Glutamate binding to the receptor's ligand-binding domains (LBDs) leads to ion channel activation and desensitization. Gating kinetics shape synaptic transmission and are strongly modulated by transmembrane AMPAR regulatory proteins (TARPs) through currently incompletely resolved mechanisms. Here, electron cryo-microscopy structures of the GluA1/2 TARP-γ8 complex, in both open and desensitized states (at 3.5 Å), reveal state-selective engagement of the LBDs by the large TARP-γ8 loop ('β1'), elucidating how this TARP stabilizes specific gating states. We further show how TARPs alter channel rectification, by interacting with the pore helix of the selectivity filter. Lastly, we reveal that the Q/R-editing site couples the channel constriction at the filter entrance to the gate, and forms the major cation binding site in the conduction path. Our results provide a mechanistic framework of how TARPs modulate AMPAR gating and conductance. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7qhh.cif.gz | 401.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7qhh.ent.gz | 280.3 KB | Display | PDB format |
| PDBx/mmJSON format | 7qhh.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/qh/7qhhftp://data.pdbj.org/pub/pdb/validation_reports/qh/7qhh | HTTPS FTP |
|---|
-Related structure data
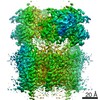
| Related structure data | 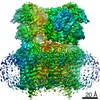 13972MC 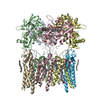 7qhbC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |

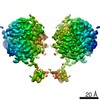
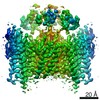
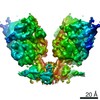
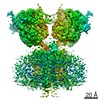





-Links
 PDBj
PDBj





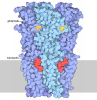
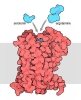


- Assembly
Assembly
| Deposited unit | 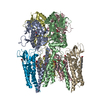
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments: Component-ID: _ / Refine code: _
NCS ensembles :
|
-Components
-Isoform Flip of Glutamate receptor ... , 2 types, 4 molecules ACBD
| #1: Protein | Mass: 102661.930 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  Homo sapiens (human) / References: UniProt: P19490 Homo sapiens (human) / References: UniProt: P19490#2: Protein | Mass: 96247.055 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / References: UniProt: P19491 |
|---|
-Protein , 1 types, 2 molecules JI
| #3: Protein | Mass: 43576.004 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / References: UniProt: Q8VHW5 |
|---|
-Non-polymers , 4 types, 18 molecules 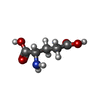
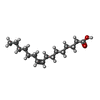
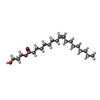
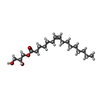
| #4: Chemical | ChemComp-GLU /  Type: L-peptide linking / Mass: 147.129 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C5H9NO4 / Feature type: SUBJECT OF INVESTIGATION Type: L-peptide linking / Mass: 147.129 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C5H9NO4 / Feature type: SUBJECT OF INVESTIGATION#5: Chemical | ChemComp-PAM /  Mass: 254.408 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C16H30O2 Mass: 254.408 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C16H30O2#6: Chemical |  Mass: 356.540 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C21H40O4 Mass: 356.540 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C21H40O4#7: Chemical |  Mass: 328.487 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C19H36O4 Mass: 328.487 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C19H36O4 |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Complex between GluA1/2 AMPA receptor and auxiliary subunit TARP gamma8 Type: COMPLEX Details: GluA2 and TARP8 are expressed as a tandem construct Entity ID: #1-#3 / Source: RECOMBINANT |
|---|---|
| Molecular weight | Value: 0.490 MDa / Experimental value: NO |
| Source (natural) | Organism: |
| Source (recombinant) | Organism: Homo sapiens (human) / Cell: HEK293 EXPI / Plasmid: PRK5 |
| Buffer solution | pH: 8 Details: 25 mM TRIS 150 mM NaCl 0.02 % GDN 87 uM CTZ 100 mM L-Glu |
| Specimen | Conc.: 2.5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: Sample was incubated with 87 uM CTZ for 30 minutes and 100 mM L-Glutamate was added before grid preparation |
| Specimen support | Grid material: COPPER / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R1.2/1.3 |
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K / Details: 3-4 second blots |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 81000 X / Nominal defocus max: 2200 nm / Nominal defocus min: 340 nm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 4 sec. / Electron dose: 51 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Num. of grids imaged: 1 / Num. of real images: 9664 |
- Processing
Processing
| Software | Name: REFMAC / Version: 5.8.0267 / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C2 (2 fold cyclic) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 105918 / Algorithm: FOURIER SPACE / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: RIGID BODY FIT / Space: REAL Details: First, rigid body fit was performed in Chimera using the coordinates of the TMD of 6QKC and the coordinates of the LBD in PDB code 3TKD. Manual model building was performed with Coot and ...Details: First, rigid body fit was performed in Chimera using the coordinates of the TMD of 6QKC and the coordinates of the LBD in PDB code 3TKD. Manual model building was performed with Coot and Refinement was performed with Refmac. The TMD and LBD maps obtained independently aided model building but were not used for refinement. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 3.6→339.2 Å / Cor.coef. Fo:Fc: 0.664 / SU B: 23.138 / SU ML: 0.306 / ESU R: 0.153 Stereochemistry target values: MAXIMUM LIKELIHOOD WITH PHASES Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Solvent model: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 81.995 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: 1 / Total: 13622 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|