 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7peq | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Model of the outer rings of the human nuclear pore complex | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | TRANSPORT PROTEIN / Nuclear Pore Complex / NPC | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationnephron development / GATOR2 complex / Seh1-associated complex / COPII-coated vesicle budding / transcription-dependent tethering of RNA polymerase II gene DNA at nuclear periphery / protein exit from endoplasmic reticulum / positive regulation of ER to Golgi vesicle-mediated transport / telomere tethering at nuclear periphery / nuclear pore complex assembly / nuclear pore outer ring ...nephron development / GATOR2 complex / Seh1-associated complex / COPII-coated vesicle budding / transcription-dependent tethering of RNA polymerase II gene DNA at nuclear periphery / protein exit from endoplasmic reticulum / positive regulation of ER to Golgi vesicle-mediated transport / telomere tethering at nuclear periphery / nuclear pore complex assembly / nuclear pore outer ring / nuclear pore organization / COPII-coated vesicle cargo loading / nuclear pore cytoplasmic filaments / COPII vesicle coat / Nuclear Pore Complex (NPC) Disassembly / Regulation of Glucokinase by Glucokinase Regulatory Protein / Defective TPR may confer susceptibility towards thyroid papillary carcinoma (TPC) / post-transcriptional tethering of RNA polymerase II gene DNA at nuclear periphery / nuclear inclusion body / Transport of Ribonucleoproteins into the Host Nucleus / positive regulation of protein exit from endoplasmic reticulum / nuclear pore nuclear basket / attachment of mitotic spindle microtubules to kinetochore / Transport of the SLBP independent Mature mRNA / Transport of the SLBP Dependant Mature mRNA / Amino acids regulate mTORC1 / SUMOylation of SUMOylation proteins / structural constituent of nuclear pore / positive regulation of mRNA splicing, via spliceosome / Transport of Mature mRNA Derived from an Intronless Transcript / NS1 Mediated Effects on Host Pathways / Rev-mediated nuclear export of HIV RNA / Nuclear import of Rev protein / SUMOylation of RNA binding proteins / macrophage chemotaxis / NEP/NS2 Interacts with the Cellular Export Machinery / RNA export from nucleus / Transport of Mature mRNA derived from an Intron-Containing Transcript / tRNA processing in the nucleus / Postmitotic nuclear pore complex (NPC) reformation / COPII-mediated vesicle transport / protein-containing complex localization / lamellipodium assembly / nucleocytoplasmic transport / nuclear localization sequence binding / Viral Messenger RNA Synthesis / poly(A)+ mRNA export from nucleus / SUMOylation of ubiquitinylation proteins / Vpr-mediated nuclear import of PICs / female gonad development / SUMOylation of DNA replication proteins / Hydrolases; Acting on peptide bonds (peptidases); Serine endopeptidases / nuclear pore / Regulation of HSF1-mediated heat shock response / positive regulation of TOR signaling / mitotic metaphase chromosome alignment / mRNA transport / SUMOylation of DNA damage response and repair proteins / mRNA export from nucleus / negative regulation of TORC1 signaling / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / positive regulation of TORC1 signaling / MHC class II antigen presentation / cellular response to nutrient levels / nuclear periphery / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / serine-type peptidase activity / cellular response to amino acid starvation / Resolution of Sister Chromatid Cohesion / SUMOylation of chromatin organization proteins / HCMV Late Events / Antigen Presentation: Folding, assembly and peptide loading of class I MHC / chromosome segregation / ER to Golgi transport vesicle membrane / Transcriptional regulation by small RNAs / molecular condensate scaffold activity / promoter-specific chromatin binding / protein import into nucleus / RHO GTPases Activate Formins / intracellular protein transport / ISG15 antiviral mechanism / spindle / kinetochore / HCMV Early Events / actin cytoskeleton / Separation of Sister Chromatids / nuclear envelope / protein transport / nuclear membrane / snRNP Assembly / ciliary basal body / nuclear body / transcription coactivator activity / ribonucleoprotein complex / cell division / lysosomal membrane / mRNA binding / endoplasmic reticulum membrane / positive regulation of DNA-templated transcription Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||
| Method | ELECTRON MICROSCOPY / subtomogram averaging / cryo EM / Resolution: 35 Å | ||||||||||||
 Authors Authors | Schuller, A.P. / Wojtynek, M. / Mankus, D. / Tatli, M. / Kronenberg-Tenga, R. / Regmi, S.G. / Dasso, M. / Weis, K. / Medalia, O. / Schwartz, T.U. | ||||||||||||
| Funding support |  Switzerland, 3items Switzerland, 3items
| ||||||||||||
 Citation Citation | Journal: Nature / Year: 2021 Title: The cellular environment shapes the nuclear pore complex architecture. Authors: Anthony P Schuller / Matthias Wojtynek / David Mankus / Meltem Tatli / Rafael Kronenberg-Tenga / Saroj G Regmi / Phat V Dip / Abigail K R Lytton-Jean / Edward J Brignole / Mary Dasso / ...Authors: Anthony P Schuller / Matthias Wojtynek / David Mankus / Meltem Tatli / Rafael Kronenberg-Tenga / Saroj G Regmi / Phat V Dip / Abigail K R Lytton-Jean / Edward J Brignole / Mary Dasso / Karsten Weis / Ohad Medalia / Thomas U Schwartz /  Abstract: Nuclear pore complexes (NPCs) create large conduits for cargo transport between the nucleus and cytoplasm across the nuclear envelope (NE). These multi-megadalton structures are composed of about ...Nuclear pore complexes (NPCs) create large conduits for cargo transport between the nucleus and cytoplasm across the nuclear envelope (NE). These multi-megadalton structures are composed of about thirty different nucleoporins that are distributed in three main substructures (the inner, cytoplasmic and nucleoplasmic rings) around the central transport channel. Here we use cryo-electron tomography on DLD-1 cells that were prepared using cryo-focused-ion-beam milling to generate a structural model for the human NPC in its native environment. We show that-compared with previous human NPC models obtained from purified NEs-the inner ring in our model is substantially wider; the volume of the central channel is increased by 75% and the nucleoplasmic and cytoplasmic rings are reorganized. Moreover, the NPC membrane exhibits asymmetry around the inner-ring complex. Using targeted degradation of Nup96, a scaffold nucleoporin of the cytoplasmic and nucleoplasmic rings, we observe the interdependence of each ring in modulating the central channel and maintaining membrane asymmetry. Our findings highlight the inherent flexibility of the NPC and suggest that the cellular environment has a considerable influence on NPC dimensions and architecture. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7peq.cif.gz | 2.5 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7peq.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 7peq.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/pe/7peqftp://data.pdbj.org/pub/pdb/validation_reports/pe/7peq | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  12814MC  7perC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data | |
| EM raw data | EMPIAR-10700 (Title: Cryo electron tomography of FIB-milled lamella of human DLD-1 cells Data size: 8.0 Data #1: Un-aligned tilt series of FIB-lamella of human DLD-1 cells [tilt series]) EMPIAR-10701 (Title: Cryo electron tomography of FIB-milled lamella of human DLD-1 cellsData size: 8.0 Data #1: Un-aligned tilt series of FIB-milled lamella of Nup96-depleted human DLD-1 cells [tilt series]) |




-Links
 PDBj
PDBj





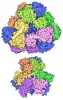



















- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 | x 8
|
-Components
-Nuclear pore complex protein ... , 5 types, 20 molecules ACBCCCDCADBDCDDDAEBECEDEAHBHCHDHAJBJCJDJ
| #1: Protein | Mass: 129108.461 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q8WUM0#2: Protein | Mass: 106504.969 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P57740#3: Protein | Mass: 106039.656 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P52948#6: Protein | Mass: 75105.266 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q9BW27#8: Protein | Mass: 162280.203 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q12769 |
|---|
-Protein , 4 types, 16 molecules AFBFCFDFAGBGCGDGAIBICIDIAKBKCKDK
| #4: Protein | Mass: 35578.438 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P55735#5: Protein | Mass: 39700.566 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q96EE3#7: Protein | Mass: 42195.652 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q8NFH3#9: Protein | Mass: 36748.512 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q8NFH4 |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: CELL / 3D reconstruction method: subtomogram averaging |
- Sample preparation
Sample preparation
| Component | Name: Nup96::Neon-AID DLD-1 / Type: CELL / Entity ID: all / Source: NATURAL |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Buffer solution | pH: 7 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: cryo-FIB milled sections of DLD1 cells |
| Specimen support | Grid material: GOLD / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R1.2/1.3 |
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE Details: Cells were grown on holey carbon, Au-mesh supports. Grids were rinsed briefly with PBS and manually blotted before plunging into liquid ethane. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 5000 nm / Nominal defocus min: 2500 nm |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Electron dose: 2.4 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING ONLY | ||||||||||||
| Symmetry | Point symmetry: C8 (8 fold cyclic) | ||||||||||||
| 3D reconstruction | Resolution: 35 Å / Resolution method: FSC 0.5 CUT-OFF / Num. of particles: 1252 / Algorithm: BACK PROJECTION / Symmetry type: POINT | ||||||||||||
| EM volume selection | Num. of tomograms: 54 / Num. of volumes extracted: 1552 | ||||||||||||
| Atomic model building | Protocol: RIGID BODY FIT | ||||||||||||
| Atomic model building | PDB-ID: 5A9Q Accession code: 5A9Q / Source name: PDB / Type: experimental model | ||||||||||||
| Refinement | Cross valid method: THROUGHOUT | ||||||||||||
| Displacement parameters | Biso max: 78.15 Å2 / Biso mean: 0.9902 Å2 / Biso min: 0 Å2 |

