 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7mpt | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Brucella melitensis NrnC with bound Mg2+ | ||||||
 要素 要素 | NanoRNase C | ||||||
 キーワード キーワード | RNA BINDING PROTEIN / RNase / bacteria / enzyme | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報 | ||||||
| 生物種 |  Brucella melitensis (マルタ熱菌) Brucella melitensis (マルタ熱菌) | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 1.75 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 1.75 Å | ||||||
 データ登録者 データ登録者 | Lormand, J.D. / Sondermann, H. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Elife / 年: 2021 タイトル: Structural characterization of NrnC identifies unifying features of dinucleotidases. 著者: Justin D Lormand / Soo-Kyoung Kim / George A Walters-Marrah / Bryce A Brownfield / J Christopher Fromme / Wade C Winkler / Jonathan R Goodson / Vincent T Lee / Holger Sondermann /  要旨: RNA degradation is fundamental for cellular homeostasis. The process is carried out by various classes of endolytic and exolytic enzymes that together degrade an RNA polymer to mono-ribonucleotides. ...RNA degradation is fundamental for cellular homeostasis. The process is carried out by various classes of endolytic and exolytic enzymes that together degrade an RNA polymer to mono-ribonucleotides. Within the exoribonucleases, nano-RNases play a unique role as they act on the smallest breakdown products and hence catalyze the final steps in the process. We recently showed that oligoribonuclease (Orn) acts as a dedicated diribonucleotidase, defining the ultimate step in RNA degradation that is crucial for cellular fitness (Kim et al., 2019). Whether such a specific activity exists in organisms that lack Orn-type exoribonucleases remained unclear. Through quantitative structure-function analyses, we show here that NrnC-type RNases share this narrow substrate length preference with Orn. Although NrnC and Orn employ similar structural features that distinguish these two classes of dinucleotidases from other exonucleases, the key determinants for dinucleotidase activity are realized through distinct structural scaffolds. The structures, together with comparative genomic analyses of the phylogeny of DEDD-type exoribonucleases, indicate convergent evolution as the mechanism of how dinucleotidase activity emerged repeatedly in various organisms. The evolutionary pressure to maintain dinucleotidase activity further underlines the important role these analogous proteins play for cell growth. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7mpt.cif.gz | 223.3 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7mpt.ent.gz | 147.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7mpt.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7mpt_validation.pdf.gz | 429 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7mpt_full_validation.pdf.gz | 429 KB | 表示 | |
| XML形式データ | 7mpt_validation.xml.gz | 22 KB | 表示 | |
| CIF形式データ | 7mpt_validation.cif.gz | 33.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/mp/7mptftp://data.pdbj.org/pub/pdb/validation_reports/mp/7mpt | HTTPS FTP |
-関連構造データ
| 関連構造データ |  7mplC  7mpmC  7mpnC  7mpoC  7mppC  7mpqC  7mprC  7mpsC  7mpuC  7mqbC  7mqcC  7mqdC  7mqeC  7mqfC  7mqgC  7mqhC  7mqiC  1yt3S S: 精密化の開始モデル C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |













-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 
| ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||
| 2 | 
| ||||||||||
| 単位格子 |
| ||||||||||
| Components on special symmetry positions |
|
-要素
| #1: タンパク質 | 分子量: 23230.486 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Brucella melitensis (マルタ熱菌)遺伝子: rnd_2, CUC12_07925, NCTC8223_00749 / 発現宿主: #2: 化合物 | ChemComp-MG /   分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg#3: 化合物 | ChemComp-PO4 / | 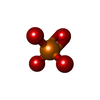  分子量: 94.971 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO4 分子量: 94.971 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO4#4: 水 | ChemComp-HOH / |  分子量: 18.015 Da / 分子数: 560 / 由来タイプ: 天然 / 式: H2O 分子量: 18.015 Da / 分子数: 560 / 由来タイプ: 天然 / 式: H2O研究の焦点であるリガンドがあるか | N | |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.9 Å3/Da / 溶媒含有率: 57.65 % |
|---|---|
| 結晶化 | 温度: 294 K / 手法: 蒸気拡散法, ハンギングドロップ法 詳細: 0.06 M sodium-potassium-monophosphate monohydrate, 1.2 M potassium phosphate dibasic, 20 % xylitol |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: APS / ビームライン: 24-ID-C / 波長: 0.9795 Å |
| 検出器 | タイプ: DECTRIS EIGER2 X 16M / 検出器: PIXEL / 日付: 2019年3月26日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.9795 Å / 相対比: 1 |
| 反射 | 解像度: 1.75→61.98 Å / Num. obs: 55765 / % possible obs: 99.77 % / 冗長度: 15.9 % / Biso Wilson estimate: 24.88 Å2 / CC1/2: 0.999 / CC star: 1 / Net I/σ(I): 26.42 |
| 反射 シェル | 解像度: 1.75→1.813 Å / 冗長度: 15 % / Mean I/σ(I) obs: 2.03 / Num. unique obs: 5452 / CC1/2: 0.754 / CC star: 0.927 / % possible all: 99.27 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 1yt3 解像度: 1.75→61.98 Å / SU ML: 0.1567 / 交差検証法: FREE R-VALUE / 位相誤差: 19.0959 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 30.35 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 1.75→61.98 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | 解像度: 1.75→1.813 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ | Refine-ID: X-RAY DIFFRACTION
|
