| 登録情報 | データベース: PDB / ID: 6ox3
|
|---|
| タイトル | SETD3 in Complex with an Actin Peptide with His73 Replaced with Lysine |
|---|
 要素 要素 | - Actin Peptide
- Histone-lysine N-methyltransferase setd3
|
|---|
 キーワード キーワード | TRANSFERASE / TRANSFERASE-STRUCTURAL PROTEIN complex |
|---|
| 機能・相同性 |  機能・相同性情報 機能・相同性情報
peptidyl-histidine methylation / regulation of uterine smooth muscle contraction / protein-histidine N-methyltransferase / protein-L-histidine N-tele-methyltransferase activity / actin modification / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex / npBAF complex / regulation of transepithelial transport ...peptidyl-histidine methylation / regulation of uterine smooth muscle contraction / protein-histidine N-methyltransferase / protein-L-histidine N-tele-methyltransferase activity / actin modification / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex / npBAF complex / regulation of transepithelial transport / nBAF complex / brahma complex / morphogenesis of a polarized epithelium / histone H3K36 methyltransferase activity / protein localization to adherens junction / postsynaptic actin cytoskeleton / structural constituent of postsynaptic actin cytoskeleton / GBAF complex / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / Tat protein binding / Gap junction degradation / regulation of G0 to G1 transition / Folding of actin by CCT/TriC / Cell-extracellular matrix interactions / dense body / regulation of nucleotide-excision repair / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / apical protein localization / regulation of double-strand break repair / adherens junction assembly / RHOF GTPase cycle / Adherens junctions interactions / histone H3K4 methyltransferase activity / tight junction / Sensory processing of sound by outer hair cells of the cochlea / Interaction between L1 and Ankyrins / SWI/SNF complex / regulation of mitotic metaphase/anaphase transition / Sensory processing of sound by inner hair cells of the cochlea / positive regulation of T cell differentiation / regulation of norepinephrine uptake / apical junction complex / transporter regulator activity / nitric-oxide synthase binding / positive regulation of muscle cell differentiation / positive regulation of double-strand break repair / maintenance of blood-brain barrier / NuA4 histone acetyltransferase complex / establishment or maintenance of cell polarity / cortical cytoskeleton / positive regulation of stem cell population maintenance / Regulation of MITF-M-dependent genes involved in pigmentation / Recycling pathway of L1 / regulation of synaptic vesicle endocytosis / regulation of G1/S transition of mitotic cell cycle / brush border / kinesin binding / EPH-ephrin mediated repulsion of cells / negative regulation of cell differentiation / RHO GTPases Activate WASPs and WAVEs / positive regulation of myoblast differentiation / RHO GTPases activate IQGAPs / positive regulation of double-strand break repair via homologous recombination / regulation of protein localization to plasma membrane / cytoskeleton organization / EPHB-mediated forward signaling / substantia nigra development / calyx of Held / axonogenesis / Translocation of SLC2A4 (GLUT4) to the plasma membrane / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / adherens junction / positive regulation of cell differentiation / actin filament / FCGR3A-mediated phagocytosis / cell motility / RHO GTPases Activate Formins / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / DNA Damage Recognition in GG-NER / Schaffer collateral - CA1 synapse / B-WICH complex positively regulates rRNA expression / kinetochore / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / PKMTs methylate histone lysines / Regulation of actin dynamics for phagocytic cup formation / structural constituent of cytoskeleton / tau protein binding / VEGFA-VEGFR2 Pathway / platelet aggregation / cytoplasmic ribonucleoprotein granule / nuclear matrix / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / UCH proteinases / Signaling by BRAF and RAF1 fusions類似検索 - 分子機能 Actin-histidine N-methyltransferase SETD3 / SETD3, SET domain / SETD3 actin-histidine N-methyltransferase (EC 2.1.1.85) family profile. / Rubisco LSMT, substrate-binding domain / Rubisco LSMT, substrate-binding domain superfamily / : / Rubisco LSMT substrate-binding / SET domain / SET domain superfamily / SET domain profile. ...Actin-histidine N-methyltransferase SETD3 / SETD3, SET domain / SETD3 actin-histidine N-methyltransferase (EC 2.1.1.85) family profile. / Rubisco LSMT, substrate-binding domain / Rubisco LSMT, substrate-binding domain superfamily / : / Rubisco LSMT substrate-binding / SET domain / SET domain superfamily / SET domain profile. / SET domain / Actins signature 1. / Actin, conserved site / Actins signature 2. / Actin/actin-like conserved site / Actins and actin-related proteins signature. / Actin / Actin family / Actin / ATPase, nucleotide binding domain類似検索 - ドメイン・相同性 ACETATE ION / TRIETHYLENE GLYCOL / S-ADENOSYL-L-HOMOCYSTEINE / Actin beta / Actin, cytoplasmic 1 / Actin-histidine N-methyltransferase類似検索 - 構成要素 |
|---|
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) |
|---|
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 1.785 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 1.785 Å |
|---|
 データ登録者 データ登録者 | Horton, J.R. / Dai, S. / Cheng, X. |
|---|
| 資金援助 |  米国, 1件 米国, 1件 | 組織 | 認可番号 | 国 |
|---|
| National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS) | GM049245-23 | 米国 |
|
|---|
 引用 引用 | #1: ジャーナル: Nature / 年: 2019タイトル: SETD3 is an actin histidine methyltransferase that prevents primary dystocia. 著者: Wilkinson, A.W. / Diep, J. / Dai, S. / Liu, S. / Ooi, Y.S. / Song, D. / Li, T.M. / Horton, J.R. / Zhang, X. / Liu, C. / Trivedi, D.V. / Ruppel, K.M. / Vilches-Moure, J.G. / Casey, K.M. / Mak, ...著者: Wilkinson, A.W. / Diep, J. / Dai, S. / Liu, S. / Ooi, Y.S. / Song, D. / Li, T.M. / Horton, J.R. / Zhang, X. / Liu, C. / Trivedi, D.V. / Ruppel, K.M. / Vilches-Moure, J.G. / Casey, K.M. / Mak, J. / Cowan, T. / Elias, J.E. / Nagamine, C.M. / Spudich, J.A. / Cheng, X. / Carette, J.E. / Gozani, O. |
|---|
| 履歴 | | 登録 | 2019年5月13日 | 登録サイト: RCSB / 処理サイト: RCSB |
|---|
| 改定 1.0 | 2019年8月21日 | Provider: repository / タイプ: Initial release |
|---|
| 改定 1.1 | 2020年1月1日 | Group: Author supporting evidence / カテゴリ: pdbx_audit_support / Item: _pdbx_audit_support.funding_organization |
|---|
| 改定 1.2 | 2023年10月11日 | Group: Data collection / Database references / Refinement description
カテゴリ: chem_comp_atom / chem_comp_bond ...chem_comp_atom / chem_comp_bond / database_2 / pdbx_initial_refinement_model
Item: _database_2.pdbx_DOI / _database_2.pdbx_database_accession |
|---|
|
|---|
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード












 PDBj
PDBj
















 集合体
集合体



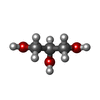

 分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H8O3
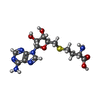
分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H8O3 タイプ: L-peptide linking / 分子量: 384.411 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C14H20N6O5S
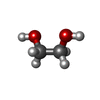
タイプ: L-peptide linking / 分子量: 384.411 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C14H20N6O5S 分子量: 62.068 Da / 分子数: 34 / 由来タイプ: 合成 / 式: C2H6O2
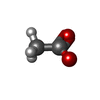
分子量: 62.068 Da / 分子数: 34 / 由来タイプ: 合成 / 式: C2H6O2 分子量: 59.044 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C2H3O2
分子量: 59.044 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C2H3O2 分子量: 150.173 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C6H14O4
分子量: 150.173 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C6H14O4 試料調製
試料調製 解析
解析