 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6hgt | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Crystal structure of human KDM4A complexed with co-substrate analog NOG and histone H3 peptide with K9R mutation | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | OXIDOREDUCTASE / Histone demethylase / Inhibitor / transcription | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報[histone H3]-trimethyl-L-lysine36 demethylase / histone H3K36me2/H3K36me3 demethylase activity / [histone H3]-trimethyl-L-lysine9 demethylase / histone H3K9me2/H3K9me3 demethylase activity / negative regulation of chromosome condensation / Barr body / histone H4K20me2 reader activity / : / histone H3K36 demethylase activity / inner kinetochore ...[histone H3]-trimethyl-L-lysine36 demethylase / histone H3K36me2/H3K36me3 demethylase activity / [histone H3]-trimethyl-L-lysine9 demethylase / histone H3K9me2/H3K9me3 demethylase activity / negative regulation of chromosome condensation / Barr body / histone H4K20me2 reader activity / : / histone H3K36 demethylase activity / inner kinetochore / pericentric heterochromatin formation / muscle cell differentiation / histone H3K9 demethylase activity / cardiac muscle hypertrophy in response to stress / oocyte maturation / histone demethylase activity / nucleosomal DNA binding / nucleus organization / pericentric heterochromatin / spermatid development / single fertilization / subtelomeric heterochromatin formation / RNA polymerase II core promoter sequence-specific DNA binding / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / NR1H3 & NR1H2 regulate gene expression linked to cholesterol transport and efflux / embryo implantation / telomere organization / Interleukin-7 signaling / Inhibition of DNA recombination at telomere / negative regulation of autophagy / RNA Polymerase I Promoter Opening / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / epigenetic regulation of gene expression / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDMs demethylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / B-WICH complex positively regulates rRNA expression / PKMTs methylate histone lysines / Meiotic recombination / Pre-NOTCH Transcription and Translation / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / multicellular organism growth / male gonad development / fibrillar center / Transcriptional regulation of granulopoiesis / HCMV Early Events / osteoblast differentiation / structural constituent of chromatin / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / nucleosome / RUNX1 regulates transcription of genes involved in differentiation of HSCs / nucleosome assembly / regulation of gene expression / HATs acetylate histones / Senescence-Associated Secretory Phenotype (SASP) / Factors involved in megakaryocyte development and platelet production / chromatin organization / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / positive regulation of cell growth / Oxidative Stress Induced Senescence / gene expression / Estrogen-dependent gene expression / chromosome, telomeric region / cell population proliferation / chromatin remodeling / cadherin binding / RNA polymerase II cis-regulatory region sequence-specific DNA binding / Amyloid fiber formation / protein heterodimerization activity / negative regulation of gene expression / negative regulation of DNA-templated transcription / ubiquitin protein ligase binding / chromatin / protein-containing complex / DNA binding / extracellular exosome / extracellular region / zinc ion binding / nucleoplasm / nucleus / membrane / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.33 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.33 Å | ||||||
 データ登録者 データ登録者 | Maw, J. / Le Bihan, Y.V. / van Montfort, R.L.M. | ||||||
| 資金援助 |  英国, 1件 英国, 1件
| ||||||
 引用 引用 | ジャーナル: To be published タイトル: Structure-based design of selective, histone substrate-based inhibitors of histone lysine demethylases 4 (KDM4) subfamily 著者: Maw, J. / Le Bihan, Y.V. / Bavetsias, V. / Blagg, J. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6hgt.cif.gz | 572.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6hgt.ent.gz | 470.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6hgt.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6hgt_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6hgt_full_validation.pdf.gz | 1.5 MB | 表示 | |
| XML形式データ | 6hgt_validation.xml.gz | 54.3 KB | 表示 | |
| CIF形式データ | 6hgt_validation.cif.gz | 76.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/hg/6hgtftp://data.pdbj.org/pub/pdb/validation_reports/hg/6hgt | HTTPS FTP |
-関連構造データ
| 関連構造データ |  2oq6S S: 精密化の開始モデル |
|---|---|
| 類似構造データ |




-リンク
 PDBj
PDBj










- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 4 | 
| ||||||||
| 単位格子 |
|
-要素
| #1: タンパク質 | 分子量: 41854.617 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: KDM4A, JHDM3A, JMJD2, JMJD2A, KIAA0677 / 発現宿主:  参照: UniProt: O75164, 酸化還元酵素; 電子対供与作用を持つ; 分子酸素を取り込むないしは分子酸素を還元する; 2- ...参照: UniProt: O75164, 酸化還元酵素; 電子対供与作用を持つ; 分子酸素を取り込むないしは分子酸素を還元する; 2-オキソグルタル酸類を片方の電子供与体とする; 酸素分をそれぞれの電子供与体に取り込む #2: タンパク質・ペプチド | 分子量: 1619.848 Da / 分子数: 4 / Mutation: K9R / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: H3F3A, H3.3A, H3F3, PP781, H3F3B, H3.3B / 発現宿主: unidentified (未定義) / 参照: UniProt: P84243, UniProt: P68431*PLUS#3: 化合物 | ChemComp-ZN /   分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#4: 化合物 | ChemComp-OGA / 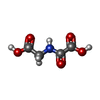  分子量: 147.086 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C4H5NO5 / タイプ: SUBJECT OF INVESTIGATION / コメント: 阻害剤*YM 分子量: 147.086 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C4H5NO5 / タイプ: SUBJECT OF INVESTIGATION / コメント: 阻害剤*YM#5: 水 | ChemComp-HOH / |  分子量: 18.015 Da / 分子数: 465 / 由来タイプ: 天然 / 式: H2O 分子量: 18.015 Da / 分子数: 465 / 由来タイプ: 天然 / 式: H2O |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.48 Å3/Da / 溶媒含有率: 50.32 % |
|---|---|
| 結晶化 | 温度: 291 K / 手法: 蒸気拡散法, シッティングドロップ法 / pH: 7.5 詳細: Crystallisation solution is 0.1M Bis-Tris-Propane pH7.5, 12-16% PEG-4000. Co-substrate analog NOG and H3R9 peptide were co-crystallised with the protein. |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Diamond / ビームライン: I04-1 / 波長: 0.9282 Å |
| 検出器 | タイプ: DECTRIS PILATUS 6M-F / 検出器: PIXEL / 日付: 2017年3月6日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.9282 Å / 相対比: 1 |
| 反射 | 解像度: 2.33→44.27 Å / Num. obs: 72206 / % possible obs: 99.7 % / 冗長度: 6.5 % / Biso Wilson estimate: 52.57 Å2 / CC1/2: 0.984 / Rmerge(I) obs: 0.18 / Net I/σ(I): 8.6 |
| 反射 シェル | 解像度: 2.33→2.39 Å / 冗長度: 7 % / Rmerge(I) obs: 1.153 / Mean I/σ(I) obs: 1.6 / Num. unique obs: 5294 / CC1/2: 0.56 / % possible all: 100 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 2oq6 解像度: 2.33→43.16 Å / Cor.coef. Fo:Fc: 0.886 / Cor.coef. Fo:Fc free: 0.86 / SU R Cruickshank DPI: 0.389 / 交差検証法: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.378 / SU Rfree Blow DPI: 0.255 / SU Rfree Cruickshank DPI: 0.26
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 44.44 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.41 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 解像度: 2.33→43.16 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | 解像度: 2.33→2.35 Å / Total num. of bins used: 50
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
