 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6dzu | ||||||
|---|---|---|---|---|---|---|---|
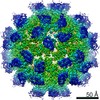
| Title | Mechanism of cellular recognition by PCV2 | ||||||
 Components Components | Putative capsid protein | ||||||
 Keywords Keywords | VIRUS LIKE PARTICLE / viral jelly-roll | ||||||
| Function / homology |  Function and homology information Function and homology informationviral capsid assembly / T=1 icosahedral viral capsid / viral penetration into host nucleus / host cell / endocytosis involved in viral entry into host cell / virion attachment to host cell / host cell nucleus / DNA binding Similarity search - Function | ||||||
| Biological species |  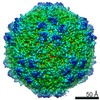 Porcine circovirus 2 Porcine circovirus 2 | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.3 Å | ||||||
| Model details | Cryo-EM image reconstruction at 3.2Angstrom resolution | ||||||
 Authors Authors | Khayat, R. / Dhindwal, S. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: J Virol / Year: 2019 Title: Porcine Circovirus 2 Uses a Multitude of Weak Binding Sites To Interact with Heparan Sulfate, and the Interactions Do Not Follow the Symmetry of the Capsid. Authors: Sonali Dhindwal / Bryant Avila / Shanshan Feng / Reza Khayat / Abstract: Porcine circovirus 2 (PCV2) is the smallest pathogenic virus capable of autonomous replication within its host. Infections result in immunosuppression and subsequent death of the host and are ...Porcine circovirus 2 (PCV2) is the smallest pathogenic virus capable of autonomous replication within its host. Infections result in immunosuppression and subsequent death of the host and are initiated via the attachment of the PCV2 icosahedral capsid to heparan sulfate (HS) and chondroitin sulfate B (CSB) glycosaminoglycans on the cell surface. However, the underlying mechanism of structural recognition remains to be explored. Using heparin, a routinely used analog of heparan sulfate, we demonstrate that increasing lengths of heparin exhibit a greater affinity toward PCV2. Our competition assays indicate that dextran sulfate (8 kDa) has a higher affinity for PCV2 than heparin (12 kDa), chondroitin sulfate B (41 kDa), hyaluronic acid (1.6 MDa), and dextran (6 kDa). This suggests that polymers high in sulfate content are capable of competing with the PCV2-heparan sulfate interaction and, thus, have the potential to inhibit PCV2 infection. Finally, we visualized the interaction between heparin and the PCV2 capsid using cryo-electron microscopy single-particle analysis, symmetry expansion, and focused classification. The image reconstructions provide the first example of an asymmetric distribution of heparin on the surface of an icosahedral virus capsid. We demonstrate that each of the 60 capsid subunits that generate the T1 capsid can bind heparin via one of five binding sites. However, not all of the binding sites were occupied by heparin, and only one-third to two-thirds of the binding sites were occupied. The binding sites are defined by arginine, lysine, and polar amino acids. Mutating the arginine, lysine, and polar amino acids to alanine diminished the binding capacity of PCV2 to heparin. It has been demonstrated that porcine circovirus 2 (PCV2) attaches to cells via heparan sulfate (HS) and chondroitin sulfate B (CSB) glycosaminoglycans; however, the underlying structural mechanism describing the HS/CSB recognition by PCV2 remains to be explored. We used cryo-electron microscopy with single-particle analysis, symmetry expansion, and focused classification to visualize the interaction between the PCV2 capsid and heparin, an analog of heparan sulfate, to better than 3.6-Å resolution. We observed that the interaction between PCV2 and heparin does not adhere to the icosahedral symmetry of the capsid. To the best of our knowledge, this is the first example where the interaction between heparin and an icosahedral capsid does not follow the symmetry elements of the capsid. Our findings also suggest that anionic polymers, such as dextran sulfate, may act to inhibit PCV2 infection. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6dzu.cif.gz | 3.5 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6dzu.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 6dzu.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/dz/6dzuftp://data.pdbj.org/pub/pdb/validation_reports/dz/6dzu | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 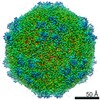 8939MC 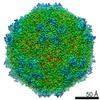 8969C 8970C 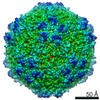 8971C 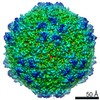 8972C 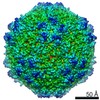 8973C 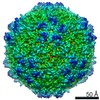 8974C 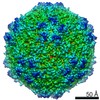 8975C  6e2rC  6e2xC  6e2zC  6e30C  6e32C  6e34C  6e39C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 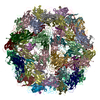
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 21913.668 Da / Num. of mol.: 60 / Fragment: Capsid protein / Mutation: None Source method: isolated from a genetically manipulated source Source: (gene. exp.) Porcine circovirus 2 / Gene: ORF2 / Plasmid: pFastBac1 / Production host:  Trichoplusia ni (cabbage looper) / Strain (production host): High Five / References: UniProt: G0ZPJ2 Trichoplusia ni (cabbage looper) / Strain (production host): High Five / References: UniProt: G0ZPJ2 |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: PCV2 / Type: VIRUS / Entity ID: all | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Units: KILODALTONS/NANOMETER / Experimental value: NO | |||||||||||||||||||||||||
| Source (natural) | Organism: Porcine circovirus 2 | |||||||||||||||||||||||||
| Details of virus | Empty: NO / Enveloped: NO / Isolate: SPECIES / Type: VIRUS-LIKE PARTICLE | |||||||||||||||||||||||||
| Natural host | Organism: Sus scrofa | |||||||||||||||||||||||||
| Virus shell | Name: Capsid protein / Diameter: 215 nm / Triangulation number (T number): 1 | |||||||||||||||||||||||||
| Buffer solution | pH: 7 | |||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||
| Specimen | Conc.: 0.718 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: This sample was monodisperse | |||||||||||||||||||||||||
| Specimen support | Details: Grids are from TedPella: 01824 / Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: Homemade | |||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 4 K |
- Electron microscopy imaging
Electron microscopy imaging
| Microscopy | Model: FEI TITAN |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Calibrated defocus min: 280 nm / Calibrated defocus max: 3200 nm / Cs: 2.7 mm / C2 aperture diameter: 100 µm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Average exposure time: 5 sec. / Electron dose: 35 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 1149 |
| Image scans | Movie frames/image: 50 / Used frames/image: 2-50 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: Per particle estimation / Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 54409 | ||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: I (icosahedral) | ||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.3 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 28938 / Algorithm: FOURIER SPACE / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||
| Atomic model building | B value: 35.5 / Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Correlation coefficient |
