 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5uym: 70S ribosome bound with cognate ternary complex base-paired to A ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5uym | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | 70S ribosome bound with cognate ternary complex base-paired to A site codon, closed 30S (Structure III) | |||||||||||||||
 Components Components |
| |||||||||||||||
 Keywords Keywords | RIBOSOME / ternary complex | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationguanyl-nucleotide exchange factor complex / protein-synthesizing GTPase / guanosine tetraphosphate binding / stringent response / negative regulation of cytoplasmic translational initiation / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / translational elongation / misfolded RNA binding / Group I intron splicing ...guanyl-nucleotide exchange factor complex / protein-synthesizing GTPase / guanosine tetraphosphate binding / stringent response / negative regulation of cytoplasmic translational initiation / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / translational elongation / misfolded RNA binding / Group I intron splicing / RNA folding / translation elongation factor activity / translational termination / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / RNA-binding transcription regulator activity / four-way junction DNA binding / negative regulation of cytoplasmic translation / DnaA-L2 complex / regulation of mRNA stability / translation repressor activity / negative regulation of translational initiation / negative regulation of DNA-templated DNA replication initiation / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / regulation of DNA-templated transcription elongation / transcription elongation factor complex / response to reactive oxygen species / cytosolic ribosome assembly / ribosome assembly / assembly of large subunit precursor of preribosome / transcription antitermination / DNA endonuclease activity / regulation of cell growth / translational initiation / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / regulation of translation / large ribosomal subunit / transferase activity / ribosomal small subunit assembly / ribosome binding / ribosome biogenesis / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / response to antibiotic / negative regulation of DNA-templated transcription / hydrolase activity / mRNA binding / GTPase activity / GTP binding / magnesium ion binding / DNA binding / RNA binding / zinc ion binding / membrane / plasma membrane / cytosol / cytoplasm Similarity search - Function | |||||||||||||||
| Biological species |  | |||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.2 Å | |||||||||||||||
 Authors Authors | Loveland, A.B. / Demo, G. / Grigorieff, N. / Korostelev, A.A. | |||||||||||||||
| Funding support |  United States, 4items United States, 4items
| |||||||||||||||
 Citation Citation | Journal: Nature / Year: 2017 Title: Ensemble cryo-EM elucidates the mechanism of translation fidelity. Authors: Anna B Loveland / Gabriel Demo / Nikolaus Grigorieff / Andrei A Korostelev / Abstract: Gene translation depends on accurate decoding of mRNA, the structural mechanism of which remains poorly understood. Ribosomes decode mRNA codons by selecting cognate aminoacyl-tRNAs delivered by ...Gene translation depends on accurate decoding of mRNA, the structural mechanism of which remains poorly understood. Ribosomes decode mRNA codons by selecting cognate aminoacyl-tRNAs delivered by elongation factor Tu (EF-Tu). Here we present high-resolution structural ensembles of ribosomes with cognate or near-cognate aminoacyl-tRNAs delivered by EF-Tu. Both cognate and near-cognate tRNA anticodons explore the aminoacyl-tRNA-binding site (A site) of an open 30S subunit, while inactive EF-Tu is separated from the 50S subunit. A transient conformation of decoding-centre nucleotide G530 stabilizes the cognate codon-anticodon helix, initiating step-wise 'latching' of the decoding centre. The resulting closure of the 30S subunit docks EF-Tu at the sarcin-ricin loop of the 50S subunit, activating EF-Tu for GTP hydrolysis and enabling accommodation of the aminoacyl-tRNA. By contrast, near-cognate complexes fail to induce the G530 latch, thus favouring open 30S pre-accommodation intermediates with inactive EF-Tu. This work reveals long-sought structural differences between the pre-accommodation of cognate and near-cognate tRNAs that elucidate the mechanism of accurate decoding. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5uym.cif.gz | 3.8 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5uym.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 5uym.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/uy/5uymftp://data.pdbj.org/pub/pdb/validation_reports/uy/5uym | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  8617MC  8615C  8616C  8618C  8619C  8620C  5uykC  5uylC  5uynC 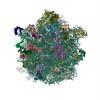 5uypC 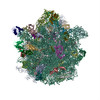 5uyqC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj






























- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
+50S ribosomal protein ... , 32 types, 32 molecules 0405060708091011121314151617181920212223242526272829303132333403
-30S ribosomal protein ... , 20 types, 20 molecules BCDEFGHIJKLMNOPQRSTU
| #32: Protein | Mass: 24253.943 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #33: Protein | Mass: 23078.785 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #34: Protein | Mass: 23383.002 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #35: Protein | Mass: 16532.088 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #36: Protein | Mass: 11669.371 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #37: Protein | Mass: 16861.523 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #38: Protein | Mass: 14015.361 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #39: Protein | Mass: 14554.882 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #40: Protein | Mass: 11196.988 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #41: Protein | Mass: 12388.068 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #42: Protein | Mass: 13636.961 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #43: Protein | Mass: 12625.753 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #44: Protein | Mass: 11475.364 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #45: Protein | Mass: 10159.621 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #46: Protein | Mass: 9207.572 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #47: Protein | Mass: 9263.946 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #48: Protein | Mass: 7606.768 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #49: Protein | Mass: 9057.626 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #50: Protein | Mass: 9506.190 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #51: Protein | Mass: 7763.073 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-RNA chain , 6 types, 7 molecules A0102XWVY
| #53: RNA chain | Mass: 498725.406 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||
|---|---|---|---|---|---|
| #54: RNA chain | Mass: 941305.250 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||
| #55: RNA chain | Mass: 38813.133 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||
| #56: RNA chain | Mass: 24802.785 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #57: RNA chain | | Mass: 5844.563 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) #58: RNA chain | | Mass: 24485.539 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Protein , 1 types, 1 molecules Z
| #59: Protein | Mass: 43152.219 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: MRE600 / Gene: tufA, b3339, JW3301 / Production host: |
|---|
-Non-polymers , 5 types, 388 molecules 

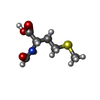
| #60: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 383 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 383 / Source method: obtained synthetically / Formula: Mg#61: Chemical |  Mass: 65.409 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Zn#62: Chemical | ChemComp-FME / |  Type: L-peptide linking / Mass: 177.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H11NO3S Type: L-peptide linking / Mass: 177.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H11NO3S#63: Chemical | ChemComp-PHE / |  Type: L-peptide linking / Mass: 165.189 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C9H11NO2 Type: L-peptide linking / Mass: 165.189 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C9H11NO2#64: Chemical | ChemComp-GCP / |  Mass: 521.208 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Formula: C11H18N5O13P3 / Comment: GMP-PCP, energy-carrying molecule analogue*YM Mass: 521.208 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Formula: C11H18N5O13P3 / Comment: GMP-PCP, energy-carrying molecule analogue*YM |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: 70S ribosome bound with cognate ternary complex base-paired to A site codon, closed 30S (Structure III) Type: RIBOSOME / Entity ID: #1-#59 / Source: MULTIPLE SOURCES | |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 2.5 MDa / Experimental value: NO | |||||||||||||||||||||||||||||||||||
| Source (natural) | Organism: | |||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | |||||||||||||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: 250 nM 50S, 250 nM 30S, 1.25 micromolar mRNA, 500 nM fMet-tRNAfMet, 1 micromolar EF-T, 500 micromolar GDPCP, 1 micromolar Phe-tRNAPhe | |||||||||||||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: C-flat-1.2/1.3 | |||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: GATAN CRYOPLUNGE 3 / Cryogen name: ETHANE / Humidity: 90 % / Chamber temperature: 275 K Details: 2 uL of complex was applied to each grid. After a 10-second incubation, the grids were blotted for 2 to 4 seconds. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 60976 X / Calibrated magnification: 60976 X / Nominal defocus max: 5000 nm / Nominal defocus min: 500 nm / Cs: 2.7 mm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 0.4 sec. / Electron dose: 1 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 2 / Num. of real images: 3928 |
| Image scans | Sampling size: 5 µm / Width: 7676 / Height: 7420 / Movie frames/image: 50 / Used frames/image: 1-50 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Image processing | Details: Gain reference was applied, movies were aligned, and the summed imaged were corrected for magnification anisotropy. | ||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Details: CTFFIND3 was used to determine CTF values. FREALIGN applied CTF correction. Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 800367 Details: Particles were picked from micrographs using Signature reference-based particle picker. | ||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.2 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 153597 / Algorithm: BACK PROJECTION / Num. of class averages: 6 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: correlation coefficient |
