 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5tc1 | ||||||
|---|---|---|---|---|---|---|---|
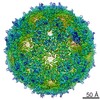
| タイトル | In situ structures of the genome and genome-delivery apparatus in ssRNA bacteriophage MS2 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | viral protein/rna / asymmetric cryoEM reconstruction / ssRNA genome structure / genome-delivery apparatus / genome-capsid interactions / viral protein-rna complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報viral genome circularization / virion attachment to host cell pilus / negative regulation of viral translation / T=3 icosahedral viral capsid / virion component / regulation of translation / structural molecule activity / RNA binding / identical protein binding 類似検索 - 分子機能 | ||||||
| 生物種 |  Enterobacteria phage MS2 (ファージ) Enterobacteria phage MS2 (ファージ) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | ||||||
 データ登録者 データ登録者 | Dai, X.H. / Li, Z.H. / Lai, M. / Shu, S. / Du, Y.S. / Zhou, Z.H. / Sun, R. | ||||||
 引用 引用 | ジャーナル: Nature / 年: 2017 タイトル: In situ structures of the genome and genome-delivery apparatus in a single-stranded RNA virus. 著者: Xinghong Dai / Zhihai Li / Mason Lai / Sara Shu / Yushen Du / Z Hong Zhou / Ren Sun /   要旨: Packaging of the genome into a protein capsid and its subsequent delivery into a host cell are two fundamental processes in the life cycle of a virus. Unlike double-stranded DNA viruses, which pump ...Packaging of the genome into a protein capsid and its subsequent delivery into a host cell are two fundamental processes in the life cycle of a virus. Unlike double-stranded DNA viruses, which pump their genome into a preformed capsid, single-stranded RNA (ssRNA) viruses, such as bacteriophage MS2, co-assemble their capsid with the genome; however, the structural basis of this co-assembly is poorly understood. MS2 infects Escherichia coli via the host 'sex pilus' (F-pilus); it was the first fully sequenced organism and is a model system for studies of translational gene regulation, RNA-protein interactions, and RNA virus assembly. Its positive-sense ssRNA genome of 3,569 bases is enclosed in a capsid with one maturation protein monomer and 89 coat protein dimers arranged in a T = 3 icosahedral lattice. The maturation protein is responsible for attaching the virus to an F-pilus and delivering the viral genome into the host during infection, but how the genome is organized and delivered is not known. Here we describe the MS2 structure at 3.6 Å resolution, determined by electron-counting cryo-electron microscopy (cryoEM) and asymmetric reconstruction. We traced approximately 80% of the backbone of the viral genome, built atomic models for 16 RNA stem-loops, and identified three conserved motifs of RNA-coat protein interactions among 15 of these stem-loops with diverse sequences. The stem-loop at the 3' end of the genome interacts extensively with the maturation protein, which, with just a six-helix bundle and a six-stranded β-sheet, forms a genome-delivery apparatus and joins 89 coat protein dimers to form a capsid. This atomic description of genome-capsid interactions in a spherical ssRNA virus provides insight into genome delivery via the host sex pilus and mechanisms underlying ssRNA-capsid co-assembly, and inspires speculation about the links between nucleoprotein complexes and the origins of viruses. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5tc1.cif.gz | 418.2 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5tc1.ent.gz | 297.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5tc1.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5tc1_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5tc1_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 5tc1_validation.xml.gz | 54.1 KB | 表示 | |
| CIF形式データ | 5tc1_validation.cif.gz | 83.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/tc/5tc1ftp://data.pdbj.org/pub/pdb/validation_reports/tc/5tc1 | HTTPS FTP |
-関連構造データ

-リンク
 PDBj
PDBj






























- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
| 詳細 | The MS2 virion contains an ssRNA chain of the genome enclosed by a capsid built from 178 copies of coat protein and a monomer of the maturation protein. Our structure shows that the maturation protein breaks the icosahedral symmetry of the capsid and imparts structural changes to some of its neighboring coat proteins (chains D, E, F, G, H in this model). All other 173 copies of the coat protein conform to the icosahedral symmetry and have same structure with the crystallographic structure of MS2 capsid with PDB ID 2MS2. Only three of these 173 chains are included in the model (chains A, B, C) for reference. |
-要素
| #1: タンパク質 | 分子量: 13869.659 Da / 分子数: 8 / 由来タイプ: 天然 詳細: The capsid of MS2 contains 178 copies of the coat protein arranged as 89 dimers in a T=3 icosahedral lattice. Structure of the capsid has been solved by crystallography with icosahedral ...詳細: The capsid of MS2 contains 178 copies of the coat protein arranged as 89 dimers in a T=3 icosahedral lattice. Structure of the capsid has been solved by crystallography with icosahedral symmetry applied (PDB ID 2MS2, also included as chains A, B, C in this model). Chains D, E, F, G, H are coat proteins that have slightly different structures in the asymmetric cryoEM reconstruction compared to the crystallographic structure. Structures of the other 170 copies of the coat protein are the same with 2MS2, and thus are not included due to the limited number of chain IDs. 由来: (天然) Enterobacteria phage MS2 (ファージ) / 参照: UniProt: P03612#2: タンパク質 | | 分子量: 44030.934 Da / 分子数: 1 / 由来タイプ: 天然 詳細: The capsid of MS2 contains a single copy of the maturation protein. Our structure shows that it replaces a coat protein dimer at one of the 2-fold icosahedral symmetry axes. Function of the ...詳細: The capsid of MS2 contains a single copy of the maturation protein. Our structure shows that it replaces a coat protein dimer at one of the 2-fold icosahedral symmetry axes. Function of the maturation protein is to attach the MS2 virion to the host (E. coli) F-pili and deliver the ssRNA viral genome into the host during infection. of the icosahedral capsid. 178 copies of the coat protein arranged as 89 dimers in a T=3 icosahedral lattice. Structure of the capsid has been solved by crystallography with icosahedral symmetry applied (PDB ID 2MS2, also included as chains A, B, C in this model). Chains D, E, F, G, H are coat proteins that have slightly different structures in the asymmetric cryoEM reconstruction compared to the crystallographic structure. Structures of the other 170 copies of the coat protein are the same with 2MS2, and thus are not included due to the limited number of chain IDs. 由来: (天然) Enterobacteria phage MS2 (ファージ) / 参照: UniProt: P03610#3: RNA鎖 | | 分子量: 1147200.750 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Enterobacteria phage MS2 (ファージ) / 参照: GenBank: 15081Has protein modification | Y | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| ||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||||||||
| ウイルスについての詳細 |
| ||||||||||||||||||||||||||||||||||||||||||
| 天然宿主 |
| ||||||||||||||||||||||||||||||||||||||||||
| ウイルス殻 |
| ||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 / 詳細: pH7.4 PBS | ||||||||||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK II / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 295 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 詳細: EFTEM mode with Gatan GIF energy filter. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / 倍率(補正後): 47170 X / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: BASIC |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最低温度: 79 K |
| 撮影 | 平均露光時間: 5.8 sec. / 電子線照射量: 50 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 2 / 実像数: 6080 |
| 電子光学装置 | エネルギーフィルター名称: GIF / エネルギーフィルター 上限: 20 eV / エネルギーフィルター 下限: 20 eV |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 7676 / 縦: 7420 / 動画フレーム数/画像: 29 / 利用したフレーム数/画像: 1-14 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 360000 | |||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 339718 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 1 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: AB INITIO MODEL / 空間: REAL |
