 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5l5y | ||||||
|---|---|---|---|---|---|---|---|
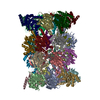
| タイトル | Yeast 20S proteasome with human beta5c (1-138) and human beta6 (97-111; 118-133) in complex with carfilzomib | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | HYDROLASE/HYDROLASE INHIBITOR / HYDROLASE-HYDROLASE INHIBITOR complex / Proteasome / Mutant / Inhibitor / Binding Analysis | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Regulation of ornithine decarboxylase (ODC) / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / proteasome core complex / proteasome core complex assembly / nuclear outer membrane-endoplasmic reticulum membrane network / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Somitogenesis ...Regulation of ornithine decarboxylase (ODC) / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / proteasome core complex / proteasome core complex assembly / nuclear outer membrane-endoplasmic reticulum membrane network / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Somitogenesis / proteasomal ubiquitin-independent protein catabolic process / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / 3-deoxy-7-phosphoheptulonate synthase / 3-deoxy-7-phosphoheptulonate synthase activity / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / KEAP1-NFE2L2 pathway / Neddylation / Orc1 removal from chromatin / MAPK6/MAPK4 signaling / proteasome storage granule / Antigen processing: Ubiquitination & Proteasome degradation / chorismate biosynthetic process / Ub-specific processing proteases / proteasome endopeptidase complex / endopeptidase activator activity / proteasome core complex, beta-subunit complex / proteasome assembly / aromatic amino acid family biosynthetic process / threonine-type endopeptidase activity / proteasome core complex, alpha-subunit complex / amino acid biosynthetic process / Regulation of activated PAK-2p34 by proteasome mediated degradation / Neutrophil degranulation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / Ubiquitin-dependent degradation of Cyclin D / proteasome complex / NIK-->noncanonical NF-kB signaling / SCF-beta-TrCP mediated degradation of Emi1 / proteolysis involved in protein catabolic process / TNFR2 non-canonical NF-kB pathway / AUF1 (hnRNP D0) binds and destabilizes mRNA / Vpu mediated degradation of CD4 / Assembly of the pre-replicative complex / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Degradation of DVL / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Dectin-1 mediated noncanonical NF-kB signaling / Degradation of AXIN / Hh mutants are degraded by ERAD / Activation of NF-kappaB in B cells / Degradation of GLI1 by the proteasome / Hedgehog ligand biogenesis / G2/M Checkpoints / Defective CFTR causes cystic fibrosis / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Autodegradation of the E3 ubiquitin ligase COP1 / Negative regulation of NOTCH4 signaling / Vif-mediated degradation of APOBEC3G / Regulation of RUNX3 expression and activity / Hedgehog 'on' state / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / MAPK6/MAPK4 signaling / Degradation of beta-catenin by the destruction complex / ABC-family proteins mediated transport / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / CDK-mediated phosphorylation and removal of Cdc6 / CLEC7A (Dectin-1) signaling / SCF(Skp2)-mediated degradation of p27/p21 / Regulation of expression of SLITs and ROBOs / FCERI mediated NF-kB activation / Regulation of PTEN stability and activity / Interleukin-1 signaling / Orc1 removal from chromatin / Regulation of RAS by GAPs / Regulation of RUNX2 expression and activity / The role of GTSE1 in G2/M progression after G2 checkpoint / Separation of Sister Chromatids / KEAP1-NFE2L2 pathway / UCH proteinases / Downstream TCR signaling / Antigen processing: Ubiquitination & Proteasome degradation / peroxisome / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Neddylation / peptidase activity / ER-Phagosome pathway / response to oxidative stress / secretory granule lumen / endopeptidase activity / proteasome-mediated ubiquitin-dependent protein catabolic process / ficolin-1-rich granule lumen / Ub-specific processing proteases / centrosome 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.7 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.7 Å | ||||||
 データ登録者 データ登録者 | Groll, M. / Huber, E.M. | ||||||
| 資金援助 |  ドイツ, 1件 ドイツ, 1件
| ||||||
 引用 引用 | ジャーナル: EMBO J. / 年: 2016 タイトル: A humanized yeast proteasome identifies unique binding modes of inhibitors for the immunosubunit beta 5i. 著者: Huber, E.M. / Heinemeyer, W. / de Bruin, G. / Overkleeft, H.S. / Groll, M. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5l5y.cif.gz | 2.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5l5y.ent.gz | 2 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5l5y.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5l5y_validation.pdf.gz | 2.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5l5y_full_validation.pdf.gz | 2.8 MB | 表示 | |
| XML形式データ | 5l5y_validation.xml.gz | 238.8 KB | 表示 | |
| CIF形式データ | 5l5y_validation.cif.gz | 306.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/l5/5l5yftp://data.pdbj.org/pub/pdb/validation_reports/l5/5l5y | HTTPS FTP |
-関連構造データ
| 関連構造データ |  5l52C  5l54C  5l55C  5l5aC  5l5bC  5l5dC  5l5eC  5l5fC  5l5hC  5l5iC  5l5jC  5l5oC  5l5pC  5l5qC  5l5rC  5l5sC  5l5tC  5l5uC  5l5vC  5l5wC  5l5xC  5l5zC  5l60C  5l61C  5l62C  5l63C  5l64C  5l65C  5l66C  5l67C  5l68C  5l69C  5l6aC  5l6bC  5l6cC  5lttC  5cz4S S: 精密化の開始モデル C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |














-リンク
 PDBj
PDBj









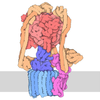





- 集合体
集合体
| 登録構造単位 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 単位格子 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 非結晶学的対称性 (NCS) | NCSドメイン:
NCSドメイン領域:
NCSアンサンブル:
NCS oper:
|
-要素
-Proteasome subunit alpha type- ... , 6種, 12分子 AOBPCQDRESGU
| #1: タンパク質 | 分子量: 27191.828 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P23639, proteasome endopeptidase complex #2: タンパク質 | 分子量: 28748.230 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P23638, proteasome endopeptidase complex #3: タンパク質 | 分子量: 28478.111 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P40303, proteasome endopeptidase complex #4: タンパク質 | 分子量: 28649.086 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P32379, proteasome endopeptidase complex #5: タンパク質 | 分子量: 25634.000 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P40302, proteasome endopeptidase complex #7: タンパク質 | 分子量: 28033.830 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P21243, proteasome endopeptidase complex |
|---|
-タンパク質 , 1種, 2分子 FT
| #6: タンパク質 | 分子量: 31575.068 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P21242, proteasome endopeptidase complex |
|---|
-Proteasome subunit beta type- ... , 7種, 14分子 HVIWJXKYLZMaNb
| #8: タンパク質 | 分子量: 25114.459 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P25043, proteasome endopeptidase complex #9: タンパク質 | 分子量: 22627.842 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P25451, proteasome endopeptidase complex #10: タンパク質 | 分子量: 22545.676 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P22141, proteasome endopeptidase complex #11: タンパク質 | 分子量: 23177.207 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Homo sapiens (ヒト), (組換発現) 遺伝子: PSMB5, LMPX, MB1, X, PRE2, DOA3, PRG1, YPR103W, P8283.10 株: ATCC 204508 / S288c / 発現宿主: 参照: UniProt: P28074, UniProt: P30656, proteasome endopeptidase complex #12: タンパク質 | 分子量: 24979.010 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Homo sapiens (ヒト), (組換発現) 株: ATCC 204508 / S288c / 遺伝子: PRE7, PRS3, PTS1, YBL041W, YBL0407, PSMB1, PSC5 / 発現宿主: 参照: UniProt: P23724, UniProt: P20618, proteasome endopeptidase complex #13: タンパク質 | 分子量: 27200.893 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P30657, proteasome endopeptidase complex #14: タンパク質 | 分子量: 21517.186 Da / 分子数: 2 / 由来タイプ: 天然 由来: (天然) 株: ATCC 204508 / S288c / 参照: UniProt: P38624, proteasome endopeptidase complex |
|---|
-非ポリマー , 5種, 419分子 

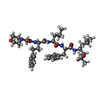
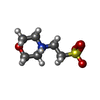

| #15: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 10 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 10 / 由来タイプ: 合成 / 式: Mg#16: 化合物 | ChemComp-CL /  分子量: 35.453 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Cl 分子量: 35.453 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Cl#17: 化合物 | ChemComp-3BV /  タイプ: peptide-like, Peptide-like / クラス: 阻害剤 / 分子量: 723.942 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C40H61N5O7 / 参照: CARFILZOMIB, bound form / コメント: 薬剤, 抗がん剤, 阻害剤*YM タイプ: peptide-like, Peptide-like / クラス: 阻害剤 / 分子量: 723.942 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C40H61N5O7 / 参照: CARFILZOMIB, bound form / コメント: 薬剤, 抗がん剤, 阻害剤*YM#18: 化合物 | ChemComp-MES /  分子量: 195.237 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C6H13NO4S / コメント: pH緩衝剤*YM 分子量: 195.237 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C6H13NO4S / コメント: pH緩衝剤*YM#19: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 395 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 3.67 Å3/Da / 溶媒含有率: 66.52 % |
|---|---|
| 結晶化 | 温度: 293 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 6.8 / 詳細: 20 mM MgAC2, 13% MPD, 0.1 M MES / PH範囲: 6.8 |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: SLS  / ビームライン: X06SA / 波長: 1 Å / ビームライン: X06SA / 波長: 1 Å |
| 検出器 | タイプ: PSI PILATUS 6M / 検出器: PIXEL / 日付: 2015年7月11日 |
| 放射 | モノクロメーター: LN2 COOLED FIXED-EXIT. SI(111) MONOCHROMATOR プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 1 Å / 相対比: 1 |
| 反射 | 解像度: 2.7→30 Å / Num. obs: 285690 / % possible obs: 97.2 % / 冗長度: 3.1 % / Rmerge(I) obs: 0.057 / Net I/σ(I): 17.1 |
| 反射 シェル | 解像度: 2.7→2.8 Å / Rmerge(I) obs: 0.474 / Mean I/σ(I) obs: 3.5 / % possible all: 98.9 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 5CZ4 解像度: 2.7→15 Å / Cor.coef. Fo:Fc: 0.953 / Cor.coef. Fo:Fc free: 0.941 / SU B: 21.711 / SU ML: 0.192 / 交差検証法: THROUGHOUT / ESU R Free: 0.247 / 立体化学のターゲット値: MAXIMUM LIKELIHOOD / 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 60.95 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.7→15 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|