 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3jck | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of the yeast 26S proteasome lid sub-complex | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | HYDROLASE / Proteasome / deubiquitinase / Rpn11 / protein homeostasis | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報SAGA complex localization to transcription regulatory region / Metalloprotease DUBs / proteasome storage granule assembly / transcription export complex 2 / protein deneddylation / peroxisome fission / maintenance of DNA trinucleotide repeats / filamentous growth / COP9 signalosome / proteasome regulatory particle ...SAGA complex localization to transcription regulatory region / Metalloprotease DUBs / proteasome storage granule assembly / transcription export complex 2 / protein deneddylation / peroxisome fission / maintenance of DNA trinucleotide repeats / filamentous growth / COP9 signalosome / proteasome regulatory particle / mitochondrial fission / proteasome regulatory particle, lid subcomplex / metal-dependent deubiquitinase activity / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / KEAP1-NFE2L2 pathway / Neddylation / proteasome binding / Orc1 removal from chromatin / MAPK6/MAPK4 signaling / regulation of protein catabolic process / proteasome storage granule / Antigen processing: Ubiquitination & Proteasome degradation / protein deubiquitination / Ub-specific processing proteases / proteasome assembly / mRNA export from nucleus / enzyme regulator activity / Neutrophil degranulation / protein folding chaperone / proteasome complex / double-strand break repair via homologous recombination / metallopeptidase activity / ubiquitin-dependent protein catabolic process / molecular adaptor activity / proteasome-mediated ubiquitin-dependent protein catabolic process / ubiquitinyl hydrolase 1 / cysteine-type deubiquitinase activity / regulation of cell cycle / structural molecule activity / positive regulation of transcription by RNA polymerase II / mitochondrion / metal ion binding / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.5 Å | ||||||
 データ登録者 データ登録者 | Herzik Jr., M.A. / Dambacher, C.M. / Worden, E.J. / Martin, A. / Lander, G.C. | ||||||
 引用 引用 | ジャーナル: Elife / 年: 2016 タイトル: Atomic structure of the 26S proteasome lid reveals the mechanism of deubiquitinase inhibition. 著者: Corey M Dambacher / Evan J Worden / Mark A Herzik / Andreas Martin / Gabriel C Lander /  要旨: The 26S proteasome is responsible for the selective, ATP-dependent degradation of polyubiquitinated cellular proteins. Removal of ubiquitin chains from targeted substrates at the proteasome is a ...The 26S proteasome is responsible for the selective, ATP-dependent degradation of polyubiquitinated cellular proteins. Removal of ubiquitin chains from targeted substrates at the proteasome is a prerequisite for substrate processing and is accomplished by Rpn11, a deubiquitinase within the 'lid' sub-complex. Prior to the lid's incorporation into the proteasome, Rpn11 deubiquitinase activity is inhibited to prevent unwarranted deubiquitination of polyubiquitinated proteins. Here we present the atomic model of the isolated lid sub-complex, as determined by cryo-electron microscopy at 3.5 Å resolution, revealing how Rpn11 is inhibited through its interaction with a neighboring lid subunit, Rpn5. Through mutagenesis of specific residues, we describe the network of interactions that are required to stabilize this inhibited state. These results provide significant insight into the intricate mechanisms of proteasome assembly, outlining the substantial conformational rearrangements that occur during incorporation of the lid into the 26S holoenzyme, which ultimately activates the deubiquitinase for substrate degradation. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3jck.cif.gz | 2.3 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3jck.ent.gz | 2 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 3jck.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3jck_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3jck_full_validation.pdf.gz | 1.9 MB | 表示 | |
| XML形式データ | 3jck_validation.xml.gz | 356.8 KB | 表示 | |
| CIF形式データ | 3jck_validation.cif.gz | 536.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/jc/3jckftp://data.pdbj.org/pub/pdb/validation_reports/jc/3jck | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj









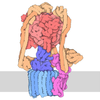





- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
| モデル数 | 5 |
-要素
-26S proteasome regulatory subunit ... , 7種, 7分子 ABCDEFH
| #1: タンパク質 | 分子量: 49573.766 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN3, SUN2, YER021W / プラスミド: pET, pCOLA, pACYC / 発現宿主:  |
|---|---|
| #2: タンパク質 | 分子量: 51840.352 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN5, NAS5, YDL147W, D1572 / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
| #3: タンパク質 | 分子量: 49839.812 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN6, NAS4, YDL097C, D2381 / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
| #4: タンパク質 | 分子量: 49016.367 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN7, YPR108W, P8283.8 / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
| #5: タンパク質 | 分子量: 38365.508 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN8, YOR261C, O5360 / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
| #6: タンパク質 | 分子量: 45839.348 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN9, NAS7, YDR427W, D9461.14 / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
| #8: タンパク質 | 分子量: 31952.119 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN12, NIN1, YFR052W / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
-タンパク質 , 2種, 2分子 GI
| #7: タンパク質 | 分子量: 34442.281 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: RPN11, MPR1, YFR004W / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
|---|---|
| #9: タンパク質 | 分子量: 10397.102 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 細胞内の位置: cytoplasm / 遺伝子: SEM1, DSH1, YDR363W-A / プラスミド: pET, pCOLA, pACYC / 発現宿主: |
-非ポリマー , 2種, 2分子 

| #10: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn |
|---|---|
| #11: 水 | ChemComp-HOH / 分子量: 18.015 Da / 分子数: 1 / 由来タイプ: 天然 / 式: H2O |
-詳細
| Has protein modification | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.37 MDa / 実験値: YES | |||||||||||||||
| 緩衝液 | 名称: 50 mM HEPES, 100 mM NaCl, 100 mM KCl, 1 mM TCEP / pH: 7.5 / 詳細: 50 mM HEPES, 100 mM NaCl, 100 mM KCl, 1 mM TCEP | |||||||||||||||
| 試料 | 濃度: 2.5 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||
| 試料支持 | 詳細: Sample was applied directly to plasma-cleaned holey carbon C-flat grids (400 mesh, 1.2 micrometer holes). | |||||||||||||||
| 急速凍結 | 装置: HOMEMADE PLUNGER / 凍結剤: ETHANE / Temp: 85 K / 湿度: 88 % 詳細: 4 uL sample was applied to the grid, blotted for 2 seconds at 4 degrees C, and plunged into liquid ethane using a manual plunger. 手法: 4 uL sample was applied to the grid, blotted for 2 seconds, and plunged into liquid ethane. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2015年2月10日 詳細: Micrograph was collected in super-resolution mode with a total frame count of 38 and total exposure time of 7.6 seconds. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 22500 X / 倍率(補正後): 38168 X / 最大 デフォーカス(公称値): 3200 nm / 最小 デフォーカス(公称値): 1600 nm / Cs: 2.7 mm 非点収差: Objective lens astigmatism was corrected at a nominal magnification of 22,500. |
| 試料ホルダ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 温度: 87.5 K / 最高温度: 90 K / 最低温度: 85 K |
| 撮影 | 電子線照射量: 43.8 e/Å2 / フィルム・検出器のモデル: GATAN K2 (4k x 4k) |
| 画像スキャン | デジタル画像の数: 3432 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: whole micrograph | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 手法: projection matching / 解像度: 3.5 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 109396 / ピクセルサイズ(公称値): 1.31 Å / ピクセルサイズ(実測値): 1.31 Å 詳細: 3D classification was performed to identify the best 109,396 particles from an initial dataset of 254,112. An ensemble of five models is provided. Each model is an approximately equivalent ...詳細: 3D classification was performed to identify the best 109,396 particles from an initial dataset of 254,112. An ensemble of five models is provided. Each model is an approximately equivalent representation of the structure. (Single particle details: Image pre-processing was performed using Appion. 3D classification and reconstruction was performed with RELION.) (Single particle--Applied symmetry: C1) 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST
|
