Herpesvirus large tegument protein deneddylase / Herpesvirus UL36 tegument protein / Herpesvirus UL35 / Herpesvirus UL35 family / Herpesvirus capsid vertex component 1 / Large tegument protein deneddylase / Herpesvirus UL17 protein / Herpesvirus tegument ubiquitin-specific protease (htUSP) domain profile. / Herpesvirus large tegument protein, USP domain / Herpesvirus tegument protein, N-terminal conserved region ...Herpesvirus large tegument protein deneddylase / Herpesvirus UL36 tegument protein / Herpesvirus UL35 / Herpesvirus UL35 family / Herpesvirus capsid vertex component 1 / Large tegument protein deneddylase / Herpesvirus UL17 protein / Herpesvirus tegument ubiquitin-specific protease (htUSP) domain profile. / Herpesvirus large tegument protein, USP domain / Herpesvirus tegument protein, N-terminal conserved region / Herpesvirus UL25 / Herpesvirus UL25 family / Herpesvirus capsid shell protein 1 / Herpesvirus capsid shell protein VP19C / Herpesvirus capsid protein 2 / Herpesvirus VP23 like capsid protein / Herpesvirus major capsid protein / Herpesvirus major capsid protein, upper domain superfamily / Herpes virus major capsid protein / Papain-like cysteine peptidase superfamily 類似検索 - ドメイン・相同性
Capsid triplex subunit 1 / UL17 / Capsid triplex subunit 2 / Major capsid protein / DNA packaging tegument protein UL25 / Large tegument protein / Small capsomere-interacting protein 類似検索 - 構成要素
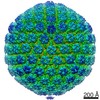
ジャーナル: mBio / 年: 2017 タイトル: The Primary Enveloped Virion of Herpes Simplex Virus 1: Its Role in Nuclear Egress. 著者: William W Newcomb / Juan Fontana / Dennis C Winkler / Naiqian Cheng / J Bernard Heymann / Alasdair C Steven / 要旨: Many viruses migrate between different cellular compartments for successive stages of assembly. The HSV-1 capsid assembles in the nucleus and then transfers into the cytoplasm. First, the capsid buds ...Many viruses migrate between different cellular compartments for successive stages of assembly. The HSV-1 capsid assembles in the nucleus and then transfers into the cytoplasm. First, the capsid buds through the inner nuclear membrane, becoming coated with nuclear egress complex (NEC) protein. This yields a primary enveloped virion (PEV) whose envelope fuses with the outer nuclear membrane, releasing the capsid into the cytoplasm. We investigated the associated molecular mechanisms by isolating PEVs from US3-null-infected cells and imaging them by cryo-electron microscopy and tomography. (pUS3 is a viral protein kinase in whose absence PEVs accumulate in the perinuclear space.) Unlike mature extracellular virions, PEVs have very few glycoprotein spikes. PEVs are ~20% smaller than mature virions, and the little space available between the capsid and the NEC layer suggests that most tegument proteins are acquired later in the egress pathway. Previous studies have proposed that NEC is organized as hexamers in honeycomb arrays in PEVs, but we find arrays of heptameric rings in extracts from US3-null-infected cells. In a PEV, NEC contacts the capsid predominantly via the pUL17/pUL25 complexes which are located close to the capsid vertices. Finally, the NEC layer dissociates from the capsid as it leaves the nucleus, possibly in response to pUS3-mediated phosphorylation. Overall, nuclear egress emerges as a process driven by a program of multiple weak interactions. On its maturation pathway, the newly formed HSV-1 nucleocapsid must traverse the nuclear envelope, while respecting the integrity of that barrier. Nucleocapsids (125 nm in diameter) are too large to pass through the nuclear pore complexes that conduct most nucleocytoplasmic traffic. It is now widely accepted that the process involves envelopment/de-envelopment of a key intermediate-the primary enveloped virion. In wild-type infections, PEVs are short-lived, which has impeded study. Using a mutant that accumulates PEVs in the perinuclear space, we were able to isolate PEVs in sufficient quantity for structural analysis by cryo-electron microscopy and tomography. The findings not only elucidate the maturation pathway of an important human pathogen but also have implications for cellular processes that involve the trafficking of large macromolecular complexes.
名称: Herpes simplex virus (type 1 / strain F) / タイプ: virus / ID: 1 / 親要素: 0 詳細: US3 null mutant, a gift from Dr. Richard Roller, Dept. of Microbiology and Immunology, Carver College of Medicine, Univ. of Iowa NCBI-ID: 10304 / 生物種: Herpes simplex virus (type 1 / strain F) / ウイルスタイプ: VIRION / ウイルス・単離状態: OTHER / ウイルス・エンベロープ: Yes / ウイルス・中空状態: No
-
実験情報
-
構造解析
手法
クライオ電子顕微鏡法
解析
単粒子再構成法
試料の集合状態
particle
-
試料調製
緩衝液
pH: 7.5 / 詳細: PBS
凍結
凍結剤: ETHANE / 装置: LEICA KF80
-
電子顕微鏡法
顕微鏡
FEI/PHILIPS CM200FEG
撮影
フィルム・検出器のモデル: GATAN ULTRASCAN 1000 (2k x 2k) 平均電子線量: 15.0 e/Å2
電子線
加速電圧: 120 kV / 電子線源: FIELD EMISSION GUN
電子光学系
照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD
+
画像解析
詳細
Images (2048 by 2048 pixels) were recorded using a CCD camera (Gatan) at a magnification of ~25,000x (giving a pixel size of 5.7 A) and processed using the Bsoft package (Heymann and Belnap, 2007) as previously described (McHugh et al., 2014).
粒子像選択
選択した数: 123 詳細: PEV images of were picked manually, yielding a total of 123 particles.
初期モデル
モデルのタイプ: OTHER 詳細: Origins and orientations were determined by projection matching, using as a starting model a map of B capsids obtained by in vitro maturation of purified procapsids (Aksyuk et al., 2015).
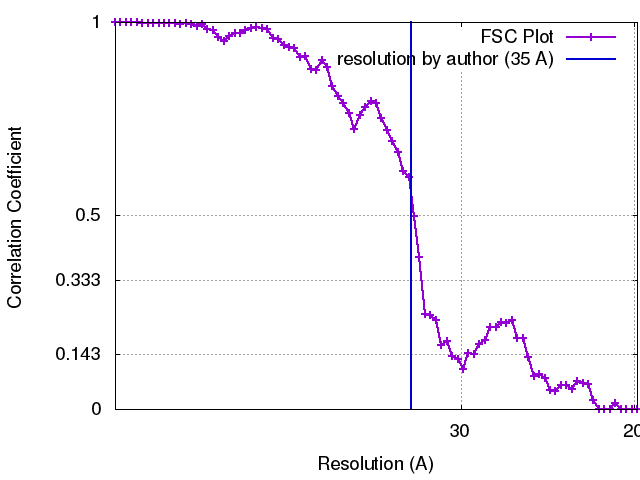
最終 再構成
想定した対称性 - 点群: I (正20面体型対称) / 解像度のタイプ: BY AUTHOR / 解像度: 35.0 Å / 解像度の算出法: FSC 0.33 CUT-OFF 詳細: Reconstructions were calculated using breconstruct (from the Bsoft package), an algorithm that integrates images as central sections of Fourier space. An inverse Fourier transform was then ...詳細: Reconstructions were calculated using breconstruct (from the Bsoft package), an algorithm that integrates images as central sections of Fourier space. An inverse Fourier transform was then calculated. Using an FSC cut-off of 0.3, the resolution of the reconstruction of C capsid-containing PEVs was estimated at ~3.5 nm for the entire particle and calculated (Cardone et al., 2013) at ~2 nm for the region around the capsid shell. 使用した粒子像数: 123
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 機能・相同性情報
機能・相同性情報 Herpes simplex virus (type 1 / strain F) (ヘルペスウイルス)
Herpes simplex virus (type 1 / strain F) (ヘルペスウイルス) データ登録者
データ登録者 引用
引用

 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク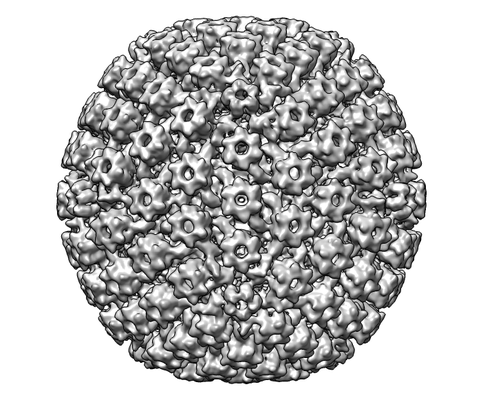 emd_8607.png
emd_8607.png http://ftp.pdbj.org/pub/emdb/structures/EMD-8607
http://ftp.pdbj.org/pub/emdb/structures/EMD-8607





 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





























 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN