 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
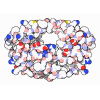
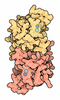
Yorodumi- EMDB-61232: Cryo-EM structure of BAF-Lamin A/C IgF-nucleosome complex (Low mo... -
+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of BAF-Lamin A/C IgF-nucleosome complex (Low mobility complex) | |||||||||||||||||||||||||||||||||
 Map data Map data | ||||||||||||||||||||||||||||||||||
 Sample Sample |
| |||||||||||||||||||||||||||||||||
 Keywords Keywords | Nucleosome / DNA binding proteins / Nuclear lamina / NUCLEAR PROTEIN | |||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationstructural constituent of nuclear lamina / negative regulation of protein ADP-ribosylation / protein localization to nuclear envelope / establishment or maintenance of microtubule cytoskeleton polarity / Breakdown of the nuclear lamina / Depolymerization of the Nuclear Lamina / nuclear envelope organization / Nuclear Envelope Breakdown / nuclear pore localization / DNA double-strand break attachment to nuclear envelope ...structural constituent of nuclear lamina / negative regulation of protein ADP-ribosylation / protein localization to nuclear envelope / establishment or maintenance of microtubule cytoskeleton polarity / Breakdown of the nuclear lamina / Depolymerization of the Nuclear Lamina / nuclear envelope organization / Nuclear Envelope Breakdown / nuclear pore localization / DNA double-strand break attachment to nuclear envelope / lamin filament / mitotic nuclear membrane reassembly / XBP1(S) activates chaperone genes / nuclear lamina / Initiation of Nuclear Envelope (NE) Reformation / Integration of viral DNA into host genomic DNA / Autointegration results in viral DNA circles / regulation of telomere maintenance / intermediate filament / nuclear migration / negative regulation of cardiac muscle hypertrophy in response to stress / muscle organ development / negative regulation of viral genome replication / 2-LTR circle formation / negative regulation of cGAS/STING signaling pathway / Vpr-mediated nuclear import of PICs / Deregulated CDK5 triggers multiple neurodegenerative pathways in Alzheimer's disease models / negative regulation of type I interferon production / Integration of provirus / APOBEC3G mediated resistance to HIV-1 infection / chromosome organization / protein localization to nucleus / negative regulation of tumor necrosis factor-mediated signaling pathway / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / condensed chromosome / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / regulation of cell migration / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / negative regulation of innate immune response / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / Interleukin-7 signaling / epigenetic regulation of gene expression / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / HCMV Late Events / SIRT1 negatively regulates rRNA expression / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDACs deacetylate histones / innate immune response in mucosa / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / lipopolysaccharide binding / Transcriptional regulation by small RNAs / HDMs demethylate histones / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / response to virus / Negative Regulation of CDH1 Gene Transcription / NoRC negatively regulates rRNA expression / G2/M DNA damage checkpoint / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / double-strand break repair via nonhomologous end joining / Meiotic recombination / Pre-NOTCH Transcription and Translation / cellular senescence / Activation of anterior HOX genes in hindbrain development during early embryogenesis / structural constituent of cytoskeleton / Transcriptional regulation of granulopoiesis / nuclear matrix / RMTs methylate histone arginines / Metalloprotease DUBs / HCMV Early Events / intracellular protein localization / structural constituent of chromatin / Signaling by BRAF and RAF1 fusions / nuclear envelope / nucleosome / UCH proteinases Similarity search - Function | |||||||||||||||||||||||||||||||||
| Biological species |  Homo sapiens (human) / synthetic construct (others) Homo sapiens (human) / synthetic construct (others) | |||||||||||||||||||||||||||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 7.14 Å | |||||||||||||||||||||||||||||||||
 Authors Authors | Horikoshi N / Miyake R / Sogawa-Fujiwara C / Ogasawara M / Takizawa Y / Kurumizaka H | |||||||||||||||||||||||||||||||||
| Funding support |  Japan, 10 items Japan, 10 items
| |||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2025 Title: Cryo-EM structures of the BAF-Lamin A/C complex bound to nucleosomes. Authors: Naoki Horikoshi / Ryosuke Miyake / Chizuru Sogawa-Fujiwara / Mitsuo Ogasawara / Yoshimasa Takizawa / Hitoshi Kurumizaka / Abstract: Barrier-to-autointegration factor (BAF) associates with mitotic chromosomes and promotes nuclear envelope assembly by recruiting proteins, such as Lamins, required for the reconstruction of the ...Barrier-to-autointegration factor (BAF) associates with mitotic chromosomes and promotes nuclear envelope assembly by recruiting proteins, such as Lamins, required for the reconstruction of the nuclear envelope and lamina. BAF also mediates chromatin anchoring to the nuclear lamina via Lamin A/C. However, the mechanism by which BAF and Lamin A/C bind chromatin and affect the chromatin organization remains elusive. Here we report the cryo-electron microscopy structures of BAF-Lamin A/C-nucleosome complexes. We find that the BAF dimer complexed with the Lamin A/C IgF domain occupies the nucleosomal dyad position, forming a tripartite nucleosomal DNA binding structure. We also show that the Lamin A/C Lys486 and His506 residues, which are reportedly mutated in lipodystrophy patients, directly contact the DNA at the nucleosomal dyad. Excess BAF-Lamin A/C complexes symmetrically bind other nucleosomal DNA sites and connect two BAF-Lamin A/C-nucleosome complexes. Although the linker histone H1 competes with BAF-Lamin A/C binding at the nucleosomal dyad region, the two BAF-Lamin A/C molecules still bridge two nucleosomes. These findings provide insights into the mechanism by which BAF, Lamin A/C, and/or histone H1 bind nucleosomes and influence chromatin organization within the nucleus. | |||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_61232.map.gz | 7.7 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-61232-v30.xmlemd-61232.xml | 27.1 KB 27.1 KB | Display Display | EMDB header |
| FSC (resolution estimation) | emd_61232_fsc.xml | 9.2 KB | Display | FSC data file |
| Images |  emd_61232.png emd_61232.png | 98.9 KB | ||
| Masks | emd_61232_msk_1.map | 64 MB | Mask map | |
| Filedesc metadata | emd-61232.cif.gz | 7.2 KB | ||
| Others | emd_61232_half_map_1.map.gzemd_61232_half_map_2.map.gz | 49.6 MB 49.6 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-61232ftp://ftp.pdbj.org/pub/emdb/structures/EMD-61232 http://ftp.pdbj.org/pub/emdb/structures/EMD-61232ftp://ftp.pdbj.org/pub/emdb/structures/EMD-61232 | HTTPS FTP |
-Related structure data
| Related structure data |  9j8nMC  9j8mC  9j8oC M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |

-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |















-Map
| File | Download / File: emd_61232.map.gz / Format: CCP4 / Size: 64 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.06 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
-Mask #1
| File | emd_61232_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








-Half map: #2
| File | emd_61232_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








-Half map: #1
| File | emd_61232_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








- Sample components
Sample components
-Entire : Low mobility complex containing BAF, Lamin A/C IgF, and nucleosome
| Entire | Name: Low mobility complex containing BAF, Lamin A/C IgF, and nucleosome |
|---|---|
| Components |
|
-Supramolecule #1: Low mobility complex containing BAF, Lamin A/C IgF, and nucleosome
| Supramolecule | Name: Low mobility complex containing BAF, Lamin A/C IgF, and nucleosome type: complex / ID: 1 / Parent: 0 / Macromolecule list: #1-#8 |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 620 KDa |
-Macromolecule #1: Histone H3.1
| Macromolecule | Name: Histone H3.1 / type: protein_or_peptide / ID: 1 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 15.719445 KDa |
| Recombinant expression | Organism:  |
| Sequence | String: GSHMARTKQT ARKSTGGKAP RKQLATKAAR KSAPATGGVK KPHRYRPGTV ALREIRRYQK STELLIRKLP FQRLVREIAQ DFKTDLRFQ SSAVMALQEA CEAYLVGLFE DTNLCAIHAK RVTIMPKDIQ LARRIRGERA UniProtKB: Histone H3.1 |
-Macromolecule #2: Histone H4
| Macromolecule | Name: Histone H4 / type: protein_or_peptide / ID: 2 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 11.676703 KDa |
| Recombinant expression | Organism: |
| Sequence | String: GSHMSGRGKG GKGLGKGGAK RHRKVLRDNI QGITKPAIRR LARRGGVKRI SGLIYEETRG VLKVFLENVI RDAVTYTEHA KRKTVTAMD VVYALKRQGR TLYGFGG UniProtKB: Histone H4 |
-Macromolecule #3: Histone H2A type 1-B/E
| Macromolecule | Name: Histone H2A type 1-B/E / type: protein_or_peptide / ID: 3 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 14.447825 KDa |
| Recombinant expression | Organism: |
| Sequence | String: GSHMSGRGKQ GGKARAKAKT RSSRAGLQFP VGRVHRLLRK GNYSERVGAG APVYLAAVLE YLTAEILELA GNAARDNKKT RIIPRHLQL AIRNDEELNK LLGRVTIAQG GVLPNIQAVL LPKKTESHHK AKGK UniProtKB: Histone H2A type 1-B/E |
-Macromolecule #4: Histone H2B type 1-J
| Macromolecule | Name: Histone H2B type 1-J / type: protein_or_peptide / ID: 4 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 14.217516 KDa |
| Recombinant expression | Organism: |
| Sequence | String: GSHMPEPAKS APAPKKGSKK AVTKAQKKDG KKRKRSRKES YSIYVYKVLK QVHPDTGISS KAMGIMNSFV NDIFERIAGE ASRLAHYNK RSTITSREIQ TAVRLLLPGE LAKHAVSEGT KAVTKYTSAK UniProtKB: Histone H2B type 1-J |
-Macromolecule #7: Barrier-to-autointegration factor
| Macromolecule | Name: Barrier-to-autointegration factor / type: protein_or_peptide / ID: 7 / Number of copies: 8 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 10.487059 KDa |
| Recombinant expression | Organism: |
| Sequence | String: GSHMMTTSQK HRDFVAEPMG EKPVGSLAGI GEVLGKKLEE RGFDKAYVVL GQFLVLKKDE DLFREWLKDT CGANAKQSRD CFGCLREWC DAFL UniProtKB: Barrier-to-autointegration factor |
-Macromolecule #8: Lamin-A/C
| Macromolecule | Name: Lamin-A/C / type: protein_or_peptide / ID: 8 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 17.551369 KDa |
| Recombinant expression | Organism: |
| Sequence | String: SHGGGSVTKK RKLESTESRS SFSQHARTSG RVAVEEVDEE GKFVRLRNKS NEDQSMGNWQ IKRQNGDDPL LTYRFPPKFT LKAGQVVTI WAAGAGATHS PPTDLVWKAQ NTWGCGNSLR TALINSTGEE VAMRKLVRSV TVVEDDEDED GDDLLHHHH UniProtKB: Prelamin-A/C |
-Macromolecule #5: DNA (193-MER)
| Macromolecule | Name: DNA (193-MER) / type: dna / ID: 5 / Number of copies: 2 / Classification: DNA |
|---|---|
| Source (natural) | Organism: synthetic construct (others) |
| Molecular weight | Theoretical: 59.351793 KDa |
| Sequence | String: (DA)(DT)(DC)(DG)(DG)(DA)(DC)(DC)(DC)(DT) (DA)(DT)(DC)(DG)(DC)(DG)(DA)(DG)(DC)(DC) (DA)(DG)(DG)(DC)(DC)(DT)(DG)(DA)(DG) (DA)(DA)(DT)(DC)(DC)(DG)(DG)(DT)(DG)(DC) (DC) (DG)(DA)(DG)(DG)(DC)(DC) ...String: (DA)(DT)(DC)(DG)(DG)(DA)(DC)(DC)(DC)(DT) (DA)(DT)(DC)(DG)(DC)(DG)(DA)(DG)(DC)(DC) (DA)(DG)(DG)(DC)(DC)(DT)(DG)(DA)(DG) (DA)(DA)(DT)(DC)(DC)(DG)(DG)(DT)(DG)(DC) (DC) (DG)(DA)(DG)(DG)(DC)(DC)(DG)(DC) (DT)(DC)(DA)(DA)(DT)(DT)(DG)(DG)(DT)(DC) (DG)(DT) (DA)(DG)(DA)(DC)(DA)(DG)(DC) (DT)(DC)(DT)(DA)(DG)(DC)(DA)(DC)(DC)(DG) (DC)(DT)(DT) (DA)(DA)(DA)(DC)(DG)(DC) (DA)(DC)(DG)(DT)(DA)(DC)(DG)(DC)(DG)(DC) (DT)(DG)(DT)(DC) (DC)(DC)(DC)(DC)(DG) (DC)(DG)(DT)(DT)(DT)(DT)(DA)(DA)(DC)(DC) (DG)(DC)(DC)(DA)(DA) (DG)(DG)(DG)(DG) (DA)(DT)(DT)(DA)(DC)(DT)(DC)(DC)(DC)(DT) (DA)(DG)(DT)(DC)(DT)(DC) (DC)(DA)(DG) (DG)(DC)(DA)(DC)(DG)(DT)(DG)(DT)(DC)(DA) (DG)(DA)(DT)(DA)(DT)(DA)(DT) (DA)(DC) (DA)(DT)(DC)(DC)(DA)(DG)(DG)(DC)(DC)(DT) (DT)(DG)(DT)(DG)(DT)(DC)(DG)(DC) (DG) (DA)(DA)(DA)(DT)(DT)(DC)(DA)(DT)(DA)(DG) (DA)(DT) |
-Macromolecule #6: DNA (193-MER)
| Macromolecule | Name: DNA (193-MER) / type: dna / ID: 6 / Number of copies: 2 / Classification: DNA |
|---|---|
| Source (natural) | Organism: synthetic construct (others) |
| Molecular weight | Theoretical: 59.823086 KDa |
| Sequence | String: (DA)(DT)(DC)(DT)(DA)(DT)(DG)(DA)(DA)(DT) (DT)(DT)(DC)(DG)(DC)(DG)(DA)(DC)(DA)(DC) (DA)(DA)(DG)(DG)(DC)(DC)(DT)(DG)(DG) (DA)(DT)(DG)(DT)(DA)(DT)(DA)(DT)(DA)(DT) (DC) (DT)(DG)(DA)(DC)(DA)(DC) ...String: (DA)(DT)(DC)(DT)(DA)(DT)(DG)(DA)(DA)(DT) (DT)(DT)(DC)(DG)(DC)(DG)(DA)(DC)(DA)(DC) (DA)(DA)(DG)(DG)(DC)(DC)(DT)(DG)(DG) (DA)(DT)(DG)(DT)(DA)(DT)(DA)(DT)(DA)(DT) (DC) (DT)(DG)(DA)(DC)(DA)(DC)(DG)(DT) (DG)(DC)(DC)(DT)(DG)(DG)(DA)(DG)(DA)(DC) (DT)(DA) (DG)(DG)(DG)(DA)(DG)(DT)(DA) (DA)(DT)(DC)(DC)(DC)(DC)(DT)(DT)(DG)(DG) (DC)(DG)(DG) (DT)(DT)(DA)(DA)(DA)(DA) (DC)(DG)(DC)(DG)(DG)(DG)(DG)(DG)(DA)(DC) (DA)(DG)(DC)(DG) (DC)(DG)(DT)(DA)(DC) (DG)(DT)(DG)(DC)(DG)(DT)(DT)(DT)(DA)(DA) (DG)(DC)(DG)(DG)(DT) (DG)(DC)(DT)(DA) (DG)(DA)(DG)(DC)(DT)(DG)(DT)(DC)(DT)(DA) (DC)(DG)(DA)(DC)(DC)(DA) (DA)(DT)(DT) (DG)(DA)(DG)(DC)(DG)(DG)(DC)(DC)(DT)(DC) (DG)(DG)(DC)(DA)(DC)(DC)(DG) (DG)(DA) (DT)(DT)(DC)(DT)(DC)(DA)(DG)(DG)(DC)(DC) (DT)(DG)(DG)(DC)(DT)(DC)(DG)(DC) (DG) (DA)(DT)(DA)(DG)(DG)(DG)(DT)(DC)(DC)(DG) (DA)(DT) |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.5 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Average electron dose: 60.1 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 2.5 µm / Nominal defocus min: 1.0 µm |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |

