endonucleolytic cleavage in 5'-ETS of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage to generate mature 5'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / (E3-independent) E2 ubiquitin-conjugating enzyme / HECT-type E3 ubiquitin transferase / nucleocytoplasmic transport / E2 ubiquitin-conjugating enzyme / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / regulation of cell size / nucleus organization / ubiquitin conjugating enzyme activity ...endonucleolytic cleavage in 5'-ETS of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage to generate mature 5'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / (E3-independent) E2 ubiquitin-conjugating enzyme / HECT-type E3 ubiquitin transferase / nucleocytoplasmic transport / E2 ubiquitin-conjugating enzyme / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / regulation of cell size / nucleus organization / ubiquitin conjugating enzyme activity / Antigen processing: Ubiquitination & Proteasome degradation / mRNA transport / protein K48-linked ubiquitination / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / protein autoubiquitination / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Neutrophil degranulation / Prevention of phagosomal-lysosomal fusion / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Membrane binding and targetting of GAG proteins / Endosomal Sorting Complex Required For Transport (ESCRT) / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / Negative regulation of FLT3 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Regulation of FZD by ubiquitination / Downregulation of ERBB4 signaling / p75NTR recruits signalling complexes / APC-Cdc20 mediated degradation of Nek2A / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Regulation of innate immune responses to cytosolic DNA / NF-kB is activated and signals survival / Downregulation of ERBB2:ERBB3 signaling / Pexophagy / NRIF signals cell death from the nucleus / VLDLR internalisation and degradation / Regulation of PTEN localization / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Regulation of BACH1 activity / MAP3K8 (TPL2)-dependent MAPK1/3 activation / TICAM1, RIP1-mediated IKK complex recruitment / Translesion synthesis by REV1 / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Translesion synthesis by POLK / InlB-mediated entry of Listeria monocytogenes into host cell / Downregulation of TGF-beta receptor signaling / Josephin domain DUBs / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / Regulation of activated PAK-2p34 by proteasome mediated degradation / Translesion synthesis by POLI / IKK complex recruitment mediated by RIP1 / Gap-filling DNA repair synthesis and ligation in GG-NER / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / TNFR1-induced NF-kappa-B signaling pathway / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / TCF dependent signaling in response to WNT / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Regulation of NF-kappa B signaling / Asymmetric localization of PCP proteins / Ubiquitin-dependent degradation of Cyclin D / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / activated TAK1 mediates p38 MAPK activation / Negative regulators of DDX58/IFIH1 signaling / TNFR2 non-canonical NF-kB pathway / AUF1 (hnRNP D0) binds and destabilizes mRNA / Regulation of signaling by CBL / NOTCH3 Activation and Transmission of Signal to the Nucleus / Vpu mediated degradation of CD4 / Assembly of the pre-replicative complex / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Degradation of DVL / Deactivation of the beta-catenin transactivating complex / Negative regulation of FGFR3 signaling / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Dectin-1 mediated noncanonical NF-kB signaling / Fanconi Anemia Pathway / Peroxisomal protein import / Degradation of AXIN / Regulation of TNFR1 signaling 類似検索 - 分子機能
E3 ubiquitin ligase, domain of unknown function DUF908 / E3 ubiquitin ligase, domain of unknown function DUF913 / Domain of Unknown Function (DUF908) / Domain of Unknown Function (DUF913) / HUWE1/Rev1, ubiquitin binding region / Ubiquitin binding region / : / HECT domain / HECT, E3 ligase catalytic domain / HECT-domain (ubiquitin-transferase) ...E3 ubiquitin ligase, domain of unknown function DUF908 / E3 ubiquitin ligase, domain of unknown function DUF913 / Domain of Unknown Function (DUF908) / Domain of Unknown Function (DUF913) / HUWE1/Rev1, ubiquitin binding region / Ubiquitin binding region / : / HECT domain / HECT, E3 ligase catalytic domain / HECT-domain (ubiquitin-transferase) / HECT domain profile. / Domain Homologous to E6-AP Carboxyl Terminus with / Ubiquitin-conjugating enzyme, active site / Ubiquitin-conjugating (UBC) active site signature. / Ubiquitin-conjugating enzyme E2 / Ubiquitin-conjugating enzyme / Ubiquitin-conjugating (UBC) core domain profile. / Ubiquitin-conjugating enzyme E2, catalytic domain homologues / Ubiquitin-conjugating enzyme/RWD-like / : / Ubiquitin domain signature. / Ubiquitin conserved site / Ubiquitin domain / Armadillo-like helical / Ubiquitin family / Ubiquitin homologues / Ubiquitin domain profile. / Ubiquitin-like domain / Armadillo-type fold / Ubiquitin-like domain superfamily 類似検索 - ドメイン・相同性
ジャーナル: Cell Rep / 年: 2025 タイトル: Structural ubiquitin contributes to K48 linkage specificity of the HECT ligase Tom1. 著者: Katrina Warner / Moritz Hunkeler / Kheewoong Baek / Anna Schmoker / Shourya S Roy Burman / Daan Overwijn / Cyrus Jin / Katherine A Donovan / Eric S Fischer / 要旨: Homologous to E6AP C terminus (HECT) ubiquitin ligases play key roles in essential pathways such as DNA repair, cell cycle control, or protein quality control. Tom1 is one of five HECT ubiquitin E3 ...Homologous to E6AP C terminus (HECT) ubiquitin ligases play key roles in essential pathways such as DNA repair, cell cycle control, or protein quality control. Tom1 is one of five HECT ubiquitin E3 ligases in budding yeast S. cerevisiae and is prototypical for a ligase with pleiotropic functions such as ubiquitin chain amplification, orphan quality control, and DNA damage response. Structures of full-length HECT ligases, including the Tom1 ortholog HUWE1, have been reported, but how domains beyond the conserved catalytic module contribute to catalysis remains largely elusive. Here, through cryoelectron microscopy (cryo-EM) snapshots of Tom1 during an active ubiquitination cycle, we demonstrate that the extended domain architecture directly contributes to activity. We identify a Tom1-ubiquitin architecture during ubiquitination involving a non-canonical ubiquitin-binding site in the solenoid shape of Tom1. We demonstrate that this ubiquitin-binding site coordinates a structural ubiquitin contributing to the fidelity of K48 poly-ubiquitin chain assembly.
超分子 #1: Class 2 Ternary complex of E3 ubiquitin ligase Tom1 with E2 ubiqu...
超分子
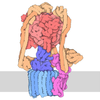
名称: Class 2 Ternary complex of E3 ubiquitin ligase Tom1 with E2 ubiquitin conjugating enzyme UBE2D2 and ubiquitin タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: all 詳細: Complex identified by plunge-freezing Tom1 in active ubiquitylation cascade
凍結剤: ETHANE / チャンバー内湿度: 90 % / チャンバー内温度: 283.15 K / 装置: LEICA PLUNGER 詳細: CHAPSO detergent added to final application at indicated concentration. Sample applied twice..
詳細
UBA1 and UBE2D2 were pre-charged for 20 minutes at room temperature in a solution containing 0.5 uM UBA1, 18 uM UBE2D2, 2 mM Mg-ATP (R and D Systems) and 25 uM ubiquitin (R and D Systems), buffered with 1X E3 Ligase Reaction Buffer (R and D Systems). Separately, Tom1 was pre-incubated with histone H2B at room temperature at room temperature for 20 minutes at a concentration of 8 uM Tom1 and 16 uM histone H2B. Samples were pre-mixed at a one-to-one volume-volume ratio for 45 seconds and 90 seconds, before addition of detergent and immediate sample application (final sample concentration: 4 uM Tom1, 8 uM histone H2B, 0.25 uM UBA1, 9 uM UBE2D2, 1 mM Mg-ATP, 12.5 uM ubiquitin, 0.2 mM CHAPSO).
-
電子顕微鏡法
顕微鏡
TFS TALOS
撮影
フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 8592 / 平均露光時間: 4.993 sec. / 平均電子線量: 52.55 e/Å2
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 米国, 2件
米国, 2件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク emd_47055.png
emd_47055.png http://ftp.pdbj.org/pub/emdb/structures/EMD-47055
http://ftp.pdbj.org/pub/emdb/structures/EMD-47055









































 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





































 試料の構成要素
試料の構成要素 Trichoplusia ni (イラクサキンウワバ)
Trichoplusia ni (イラクサキンウワバ)
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
