 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM map of full-length Tom1 (S. cerevisiae), HECT focus map | |||||||||
 マップデータ マップデータ | consensus, Tom1 BS3 crosslink HECT domain focus map | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | transferase / ubiquitin / ubiquitylation / complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報endonucleolytic cleavage in 5'-ETS of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage to generate mature 5'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / HECT-type E3 ubiquitin transferase / nucleocytoplasmic transport / regulation of cell size / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / nucleus organization / Antigen processing: Ubiquitination & Proteasome degradation / mRNA transport / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) ...endonucleolytic cleavage in 5'-ETS of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage to generate mature 5'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / HECT-type E3 ubiquitin transferase / nucleocytoplasmic transport / regulation of cell size / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / nucleus organization / Antigen processing: Ubiquitination & Proteasome degradation / mRNA transport / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / Neutrophil degranulation / ubiquitin-protein transferase activity / ubiquitin protein ligase activity / mitotic cell cycle / ubiquitin-dependent protein catabolic process / protein ubiquitination / nucleolus / nucleus / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.2 Å | |||||||||
 データ登録者 データ登録者 | Warner KM / Hunkeler M / Fischer ES | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Cell Rep / 年: 2025 タイトル: Structural ubiquitin contributes to K48 linkage specificity of the HECT ligase Tom1. 著者: Katrina Warner / Moritz Hunkeler / Kheewoong Baek / Anna Schmoker / Shourya S Roy Burman / Daan Overwijn / Cyrus Jin / Katherine A Donovan / Eric S Fischer / 要旨: Homologous to E6AP C terminus (HECT) ubiquitin ligases play key roles in essential pathways such as DNA repair, cell cycle control, or protein quality control. Tom1 is one of five HECT ubiquitin E3 ...Homologous to E6AP C terminus (HECT) ubiquitin ligases play key roles in essential pathways such as DNA repair, cell cycle control, or protein quality control. Tom1 is one of five HECT ubiquitin E3 ligases in budding yeast S. cerevisiae and is prototypical for a ligase with pleiotropic functions such as ubiquitin chain amplification, orphan quality control, and DNA damage response. Structures of full-length HECT ligases, including the Tom1 ortholog HUWE1, have been reported, but how domains beyond the conserved catalytic module contribute to catalysis remains largely elusive. Here, through cryoelectron microscopy (cryo-EM) snapshots of Tom1 during an active ubiquitination cycle, we demonstrate that the extended domain architecture directly contributes to activity. We identify a Tom1-ubiquitin architecture during ubiquitination involving a non-canonical ubiquitin-binding site in the solenoid shape of Tom1. We demonstrate that this ubiquitin-binding site coordinates a structural ubiquitin contributing to the fidelity of K48 poly-ubiquitin chain assembly. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_47035.map.gz | 203.9 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-47035-v30.xmlemd-47035.xml | 24.2 KB 24.2 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_47035_fsc.xml | 12.7 KB | 表示 | FSCデータファイル |
| 画像 |  emd_47035.png emd_47035.png | 75.9 KB | ||
| マスクデータ | emd_47035_msk_1.mapemd_47035_msk_2.map | 216 MB 216 MB | マスクマップ | |
| Filedesc metadata | emd-47035.cif.gz | 7.9 KB | ||
| その他 | emd_47035_additional_1.map.gzemd_47035_half_map_1.map.gzemd_47035_half_map_2.map.gz | 188.9 MB 200.2 MB 200.2 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-47035ftp://ftp.pdbj.org/pub/emdb/structures/EMD-47035 http://ftp.pdbj.org/pub/emdb/structures/EMD-47035ftp://ftp.pdbj.org/pub/emdb/structures/EMD-47035 | HTTPS FTP |
-関連構造データ
















-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |





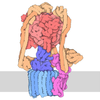





-マップ
| ファイル | ダウンロード / ファイル: emd_47035.map.gz / 形式: CCP4 / 大きさ: 216 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | consensus, Tom1 BS3 crosslink HECT domain focus map | ||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 0.83 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
-マスク #1
| ファイル | emd_47035_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-マスク #2
| ファイル | emd_47035_msk_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-追加マップ: DeepEMhancer map, Tom1 BS3 crosslink HECT domain focus map
| ファイル | emd_47035_additional_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | DeepEMhancer map, Tom1 BS3 crosslink HECT domain focus map | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: half map A, Tom1 BS3 crosslink HECT domain focus map
| ファイル | emd_47035_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | half map A, Tom1 BS3 crosslink HECT domain focus map | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: half map B, Tom1 BS3 crosslink HECT domain focus map
| ファイル | emd_47035_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | half map B, Tom1 BS3 crosslink HECT domain focus map | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








- 試料の構成要素
試料の構成要素
-全体 : Full length structure, S. cerevisiae Tom1
| 全体 | 名称: Full length structure, S. cerevisiae Tom1 |
|---|---|
| 要素 |
|
-超分子 #1: Full length structure, S. cerevisiae Tom1
| 超分子 | 名称: Full length structure, S. cerevisiae Tom1 / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: all / 詳細: Tom1 closed-ring conformation, stabilized |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 376 KDa |
-分子 #1: E3 ubiquitin-protein ligase Tom1
| 分子 | 名称: E3 ubiquitin-protein ligase Tom1 / タイプ: protein_or_peptide / ID: 1 / 詳細: StrepIItag-TEV-Tom1 fusion / 光学異性体: LEVO / EC番号: HECT-type E3 ubiquitin transferase |
|---|---|
| 由来(天然) | 生物種: |
| 組換発現 | 生物種:  Trichoplusia ni (イラクサキンウワバ) Trichoplusia ni (イラクサキンウワバ) |
| 配列 | 文字列: MDWSHPQFEK SAVDENLYFQ GGGRMVLFTR CEKARKEKLA AGYKPLVDYL IDCDTPTFLE RIEAIQEWDR SRDDLYVWIP ILDRMDGLLL KVAEKYKYKQ DPKKECEVKL VEMEAHDVDY CLKMLKFTRR LLLNTENRFV YSSGDVLMYL LNCPNFTIKL AVMRILAILG ...文字列: MDWSHPQFEK SAVDENLYFQ GGGRMVLFTR CEKARKEKLA AGYKPLVDYL IDCDTPTFLE RIEAIQEWDR SRDDLYVWIP ILDRMDGLLL KVAEKYKYKQ DPKKECEVKL VEMEAHDVDY CLKMLKFTRR LLLNTENRFV YSSGDVLMYL LNCPNFTIKL AVMRILAILG ERFVIAREKI VAHNIFGDHN LRKKTLKLAL SLSSSVMDED GEHFSLVDLY FDKKKVPQKW RKLRFTHYTS NDFKKSSQQK NNINETQTSI KKVTMTTQEL CEHSLQQIFD KGMALLPAES WFDFSIKASV AKAFSDDSGE NIDLRNIIIE TKLNAIAFVN TIFSPPQVSS KLFELDPYAF NSLTDLISLS ETKIPKELRT DALFTLECIS LKHVWCSDII RNLGGNISHG LLFQILRYIA KTLREATDEI DEEYNVRFFY LISNLADVKP LHESLFAAGL IPTLLEIVSI RNCPYKRTLA SATHLLETFI DNSETTTEFI ENDGFTMLIT SVANEIDFTL AHPETWQPPK YSVVYYSISF RELAYIRSLL KLVLKLLSTD SGDRIRNLID SPILVSLKKI LENKLVFGLT LITYTLDVVQ KVINSEPTIY PVLVEAGLIP YVIDNFPKLI GPSAELLSLL PDVVSAICLN PEGLKQVKEK GLINNLFDFL LDADHARILT GGDRSTEYGT DIDELARHYP DLKANIVEAL CNVIRKMPST FRNEREFLFT SPKDQKYFFH RKNEEILTDK EEHEPAYWEL LDKGTMLDTF TSVLFGMSLG NGSFSQVPQH LEARDFLAII FMENPPYEYF TSVAISNVTE VLQYLDEKYE DYAFMDVMKV LNDQLENLND FLNSPNDRSF FLERDGENSV RSCHSKLCRL AAILNIVTNV YIDLTTLSCK RIMQIYSYFD KRGFSLIKNL KLLFQKCALE EMYIRQHMPD SVITETMPLP IVDVSGDGPP LQIYIDDPKK GDQKGKITSV KTRNTLQMRT ILYTLQSNTA ILFRCFLRLS HSRNMDLEHK DLTTEVHIFE NVVENVIEML KATELEGHLP YILVLLNFNT FVFTIPKASP NSTEILQTIP AYIFYQKGGY LLYLHIIRDL FTRMTKIKDL SSLDNINYID ESNGILTLSC LINALTFYNK SMQTETMENV QSIGKYYVSI DDDYNIMKAL TVPIKVMALA MILDLDKSDS LFKTQSRNVP YSVFKQLLSM LKNIFTNVNI YTKELYELHW DLIFPPIKKI SLFEQVGIPG DVAANYLTDT GDDLPADNSI GLFSPEQWEK YKKLIGEDKS IYYPQPMQAQ YYKGCSSKEL DELRDTFFND GLPSRIFTVL PFYPKLVNAF AKTLLQIFTK YDEPTEVFAG RILDRILETD LDDPATLSSL IHLFGIFLNE KYIYQKASHL MQRFIEYLEK SLKPEHVNTP WFSKALYVYE IILAKSELPH LEELSKDVLL RYPLLSMAKV FRIPDPMKQK LFDILIRVSD ISNFYSALAT SRILIFYSRD ELYANNIARS GILSRLLKVI GSFQKLDKIN FLESSFLLLT RRCFETTENV DALIRAEINR SFTARPLGGG DDAVRELTTI LEEKAHVVMR SPSQFIDVLC ETARFHEFDD QGALVDYSLK RFLGEKDKNT QASSTEKSDI YERTGIMHLL LSQLMAASEK DWLSEPANSS DLPENKKAQL DPSRNPVCAY MIFLLKLLVE LVSSYNQCKF EFLTFSRRNT YAERPRPRTT AINFFLYRLL DKPVGTDHDK HEAKRREVIG MLARSVIIGF LATVQDDRTT KTDVKLADPH MNFIRKFAIE AIIKAIRNAT SSSKLLESNH LKLDMWFRII TSMVYVQAPY LRQLLDSNKV EADQYQLCKL VIDLGLPSVI TEAMASIDLN YPFSKKIFNV AVEALNTISS TRNNFSEHFK IEDHDEVEDE VDESDKEEIP DMFKNSALGM YDVEDIEEDD DDDTSLIGDD DAMAFVDSDN GFEVVFSDED DDMGEEDADD ARSDSEENEL SSEMQSSTAD GTDVDYEVDD ADGLIINIDQ PSGDDEEMAD YDANISHSSH SENEDDASMD VIEVYDDELS SGYDVDLSDY DVDESDWDSG LSSLSISDED SESSEDEPIN STRMGDSRRR WLIAEGVELT DDSQGESEED DRGVFRGIEH IFSNENEPLF RVHDEMRHRN HHRSINRTHF HSAMSAPSLS LLNRGRRNQS NLINPLGPTG LEQVENDISD QVTVAGSGSR PRSHHLHFSE VLVSGSFFDE PVLDGIILKS TVSRWKDIFD MFYDSKTYAN CIIPTVINRL YKVSLALQKD LENKREQEKL KNKNLLFNEA KVESHNSSDA ISVEQDDIQE SNVTHDDHEP VYVTIQGSEV DIGGTDIDPE FMNALPDDIR ADVFAQHVRE RRAEARLNSD HNVHSREIDS DFLEAIPEDI REGILDTEAE EQRMFGRIGS SADVIRADDD VSNNDEEVEN GLDHGNSNDR NNADPEKKKP ARIYFAPLID RAGIASLMKS VFISKPYIQR EIYHELFYRL CSSKQNRNDL MNTFLFILSE GIIDQHSLEK VYNIISSRAM GHAKTTTVRQ LPSDCTPLTV ANQTIEILQS LIDADSRLKY FLIAEHDNLI VNKANNKSRK EALPDKKLRW PLWHLFSLLD RKLITDESVL MDLLTRILQV CTKTLAVLST SSNGKENLSK KFHLPSFDED DLMKILSIIM LDSCTTRVFQ QTLNIIYNLS KLQGCMSIFT KHLVSLAISI MSKLKSALDG LSREVGTITT GMEINSELLQ KFTLPSSDQA KLLKILTTVD FLYTHKRKEE ERNVKDLQSL YDKMNGGPVW SSLSECLSQF EKSQAINTSA TILLPLIESL MVVCRRSDLS QNRNTAVKYE DAKLLDFSKT RVENLFFPFT DAHKKLLNQM IRSNPKLMSG PFALLVKNPK VLDFDNKRYF FNAKLKSDNQ ERPKLPITVR REQVFLDSYR ALFFKTNDEI KNSKLEITFK GESGVDAGGV TREWYQVLSR QMFNPDYALF LPVPSDKTTF HPNRTSGINP EHLSFFKFIG MIIGKAIRDQ CFLDCHFSRE VYKNILGRPV SLKDMESLDP DYYKSLVWIL ENDITDIIEE TFSVETDDYG EHKVINLIEG GKDIIVTEAN KQDYVKKVVE YKLQTSVKEQ MDNFLVGFYA LISKDLITIF DEQELELLIS GLPDIDVDDW KNNTTYVNYT ATCKEVSYFW RAVRSFDAEE RAKLLQFVTG TSKVPLNGFK ELSGVNGVCK FSIHRDFGSS ERLPSSHTCF NQLNLPPYES YETLRGSLLL AINEGHEGFG LA UniProtKB: E3 ubiquitin-protein ligase TOM1 |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 4.8 mg/mL | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | pH: 7.2 構成要素:
詳細: 30 mM HEPES pH 7.2, 200 mM NaCl, 2 mM TCEP, 0.3 mM CHAPSO | |||||||||||||||
| グリッド | モデル: Quantifoil R1.2/1.3 / 材質: COPPER / メッシュ: 300 / 支持フィルム - 材質: CARBON / 支持フィルム - トポロジー: HOLEY ARRAY / 前処理 - タイプ: GLOW DISCHARGE / 前処理 - 時間: 60 sec. / 前処理 - 雰囲気: AIR / 前処理 - 気圧: 0.039 kPa / 詳細: 60 seconds, hold time 10 seconds, Current 15 mA | |||||||||||||||
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 90 % / チャンバー内温度: 283 K / 装置: LEICA EM GP 詳細: CHAPSO detergent added to final conc. of 0.3 mM. Sample applied twice.. |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | TFS KRIOS |
|---|---|
| 特殊光学系 | エネルギーフィルター - 名称: GIF Bioquantum / エネルギーフィルター - スリット幅: 20 eV |
| 撮影 | フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 11583 / 平均露光時間: 2.8 sec. / 平均電子線量: 53.687 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 50.0 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 2.2 µm / 最小 デフォーカス(公称値): 1.1 µm / 倍率(公称値): 105000 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
+画像解析

-原子モデル構築 1
| 初期モデル | PDB ID: Chain - Source name: AlphaFold / Chain - Initial model type: in silico model |
|---|

