 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | biogenesis module of human telomerase holoenzyme | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | Telomerase / REPLICATION | |||||||||
| Function / homology |  Function and homology information Function and homology informationtelomere formation via telomerase / box H/ACA scaRNP complex / box H/ACA telomerase RNP complex / protein localization to Cajal body / snoRNA guided rRNA pseudouridine synthesis / enzyme-directed rRNA pseudouridine synthesis / pseudouridine synthesis / Cajal body organization / rRNA pseudouridine synthesis / Isomerases; Intramolecular transferases; Transferring other groups ...telomere formation via telomerase / box H/ACA scaRNP complex / box H/ACA telomerase RNP complex / protein localization to Cajal body / snoRNA guided rRNA pseudouridine synthesis / enzyme-directed rRNA pseudouridine synthesis / pseudouridine synthesis / Cajal body organization / rRNA pseudouridine synthesis / Isomerases; Intramolecular transferases; Transferring other groups / box H/ACA sno(s)RNA 3'-end processing / box H/ACA snoRNP complex / telomerase RNA stabilization / regulation of telomerase RNA localization to Cajal body / protein carrier activity / snRNA pseudouridine synthesis / mRNA pseudouridine synthesis / box H/ACA snoRNA binding / pseudouridine synthase activity / telomerase RNA localization to Cajal body / telomerase activity / positive regulation of establishment of protein localization to telomere / scaRNA localization to Cajal body / positive regulation of telomerase RNA localization to Cajal body / RNA folding chaperone / sno(s)RNA-containing ribonucleoprotein complex / telomerase holoenzyme complex / telomerase RNA binding / U3 snoRNA binding / positive regulation of double-strand break repair via nonhomologous end joining / rRNA modification in the nucleus and cytosol / telomerase holoenzyme complex assembly / positive regulation of double-strand break repair / Association of TriC/CCT with target proteins during biosynthesis / Telomere Extension By Telomerase / RNA folding / Cajal body / telomere maintenance via telomerase / RNA processing / positive regulation of double-strand break repair via homologous recombination / positive regulation of telomere maintenance via telomerase / positive regulation of DNA repair / mRNA 3'-UTR binding / fibrillar center / rRNA processing / site of double-strand break / protein-folding chaperone binding / histone binding / chromosome, telomeric region / nuclear body / DNA repair / ubiquitin protein ligase binding / protein-containing complex binding / nucleolus / RNA binding / nucleoplasm / identical protein binding / nucleus / cytoplasm / cytosol Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.94 Å | |||||||||
 Authors Authors | Wan F / Ding Y | |||||||||
| Funding support |  China, 1 items China, 1 items
| |||||||||
 Citation Citation | Journal: Cell Res / Year: 2021 Title: Zipper head mechanism of telomere synthesis by human telomerase. Authors: Futang Wan / Yongbo Ding / Yuebin Zhang / Zhenfang Wu / Shaobai Li / Lin Yang / Xiangyu Yan / Pengfei Lan / Guohui Li / Jian Wu / Ming Lei / Abstract: Telomerase, a multi-subunit ribonucleoprotein complex, is a unique reverse transcriptase that catalyzes the processive addition of a repeat sequence to extend the telomere end using a short fragment ...Telomerase, a multi-subunit ribonucleoprotein complex, is a unique reverse transcriptase that catalyzes the processive addition of a repeat sequence to extend the telomere end using a short fragment of its own RNA component as the template. Despite recent structural characterizations of human and Tetrahymena telomerase, it is still a mystery how telomerase repeatedly uses its RNA template to synthesize telomeric DNA. Here, we report the cryo-EM structure of human telomerase holoenzyme bound with telomeric DNA at resolutions of 3.5 Å and 3.9 Å for the catalytic core and biogenesis module, respectively. The structure reveals that a leucine residue Leu980 in telomerase reverse transcriptase (TERT) catalytic subunit functions as a zipper head to limit the length of the short primer-template duplex in the active center. Moreover, our structural and computational analyses suggest that TERT and telomerase RNA (hTR) are organized to harbor a preformed active site that can accommodate short primer-template duplex substrates for catalysis. Furthermore, our findings unveil a double-fingers architecture in TERT that ensures nucleotide addition processivity of human telomerase. We propose that the zipper head Leu980 is a structural determinant for the sequence-based pausing signal of DNA synthesis that coincides with the RNA element-based physical template boundary. Functional analyses unveil that the non-glycine zipper head plays an essential role in both telomerase repeat addition processivity and telomere length homeostasis. In addition, we also demonstrate that this zipper head mechanism is conserved in all eukaryotic telomerases. Together, our study provides an integrated model for telomerase-mediated telomere synthesis. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_31813.map.gz | 257.7 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-31813-v30.xmlemd-31813.xml | 14.8 KB 14.8 KB | Display Display | EMDB header |
| Images | 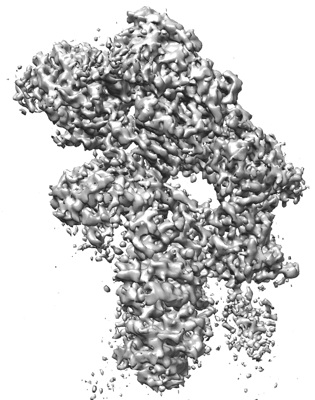 emd_31813.png emd_31813.png | 120.4 KB | ||
| Filedesc metadata | emd-31813.cif.gz | 6 KB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-31813ftp://ftp.pdbj.org/pub/emdb/structures/EMD-31813 http://ftp.pdbj.org/pub/emdb/structures/EMD-31813ftp://ftp.pdbj.org/pub/emdb/structures/EMD-31813 | HTTPS FTP |
-Related structure data
| Related structure data |  7v9aMC  7v99C C: citing same article ( M: atomic model generated by this map |
|---|---|
| Similar structure data |



-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |












-Map
| File | Download / File: emd_31813.map.gz / Format: CCP4 / Size: 274.6 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.1 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
- Sample components
Sample components
-Entire : Biogenesis module
| Entire | Name: Biogenesis module |
|---|---|
| Components |
|
-Supramolecule #1: Biogenesis module
| Supramolecule | Name: Biogenesis module / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
-Macromolecule #1: Telomerase Cajal body protein 1
| Macromolecule | Name: Telomerase Cajal body protein 1 / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 59.35707 KDa |
| Sequence | String: MKTLETQPLA PDCCPSDQDP APAHPSPHAS PMNKNADSEL MPPPPERGDP PRLSPDPVAG SAVSQELREG DPVSLSTPLE TEFGSPSEL SPRIEEQELS ENTSLPAEEA NGSLSEEEAN GPELGSGKAM EDTSGEPAAE DEGDTAWNYS FSQLPRFLSG S WSEFSTQP ...String: MKTLETQPLA PDCCPSDQDP APAHPSPHAS PMNKNADSEL MPPPPERGDP PRLSPDPVAG SAVSQELREG DPVSLSTPLE TEFGSPSEL SPRIEEQELS ENTSLPAEEA NGSLSEEEAN GPELGSGKAM EDTSGEPAAE DEGDTAWNYS FSQLPRFLSG S WSEFSTQP ENFLKGCKWA PDGSCILTNS ADNILRIYNL PPELYHEGEQ VEYAEMVPVL RMVEGDTIYD YCWYSLMSSA QP DTSYVAS SSRENPIHIW DAFTGELRAS FRAYNHLDEL TAAHSLCFSP DGSQLFCGFN RTVRVFSTAR PGRDCEVRAT FAK KQGQSG IISCIAFSPA QPLYACGSYG RSLGLYAWDD GSPLALLGGH QGGITHLCFH PDGNRFFSGA RKDAELLCWD LRQS GYPLW SLGREVTTNQ RIYFDLDPTG QFLVSGSTSG AVSVWDTDGP GNDGKPEPVL SFLPQKDCTN GVSLHPSLPL LATAS GQRV FPEPTESGDE GEELGLPLLS TRHVHLECRL QLWWCGGAPD SSIPDDHQGE KGQGGTEGGV GELI UniProtKB: Telomerase Cajal body protein 1 |
-Macromolecule #2: H/ACA ribonucleoprotein complex subunit DKC1
| Macromolecule | Name: H/ACA ribonucleoprotein complex subunit DKC1 / type: protein_or_peptide / ID: 2 / Number of copies: 2 / Enantiomer: LEVO EC number: Isomerases; Intramolecular transferases; Transferring other groups |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 57.779211 KDa |
| Sequence | String: MADAEVIILP KKHKKKKERK SLPEEDVAEI QHAEEFLIKP ESKVAKLDTS QWPLLLKNFD KLNVRTTHYT PLACGSNPLK REIGDYIRT GFINLDKPSN PSSHEVVAWI RRILRVEKTG HSGTLDPKVT GCLIVCIERA TRLVKSQQSA GKEYVGIVRL H NAIEGGTQ ...String: MADAEVIILP KKHKKKKERK SLPEEDVAEI QHAEEFLIKP ESKVAKLDTS QWPLLLKNFD KLNVRTTHYT PLACGSNPLK REIGDYIRT GFINLDKPSN PSSHEVVAWI RRILRVEKTG HSGTLDPKVT GCLIVCIERA TRLVKSQQSA GKEYVGIVRL H NAIEGGTQ LSRALETLTG ALFQRPPLIA AVKRQLRVRT IYESKMIEYD PERRLGIFWV SCEAGTYIRT LCVHLGLLLG VG GQMQELR RVRSGVMSEK DHMVTMHDVL DAQWLYDNHK DESYLRRVVY PLEKLLTSHK RLVMKDSAVN AICYGAKIML PGV LRYEDG IEVNQEIVVI TTKGEAICMA IALMTTAVIS TCDHGIVAKI KRVIMERDTY PRKWGLGPKA SQKKLMIKQG LLDK HGKPT DSTPATWKQE YVDYSESAKK EVVAEVVKAP QVVAEAAKTA KRKRESESES DETPPAAPQL IKKEKKKSKK DKKAK AGLE SGAEPGDGDS DTTKKKKKKK KAKEVELVSE UniProtKB: H/ACA ribonucleoprotein complex subunit DKC1 |
-Macromolecule #3: H/ACA ribonucleoprotein complex subunit 1
| Macromolecule | Name: H/ACA ribonucleoprotein complex subunit 1 / type: protein_or_peptide / ID: 3 / Number of copies: 2 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 22.387963 KDa |
| Sequence | String: MSFRGGGRGG FNRGGGGGGF NRGGSSNHFR GGGGGGGGGN FRGGGRGGFG RGGGRGGFNK GQDQGPPERV VLLGEFLHPC EDDIVCKCT TDENKVPYFN APVYLENKEQ IGKVDEIFGQ LRDFYFSVKL SENMKASSFK KLQKFYIDPY KLLPLQRFLP R PPGEKGPP ...String: MSFRGGGRGG FNRGGGGGGF NRGGSSNHFR GGGGGGGGGN FRGGGRGGFG RGGGRGGFNK GQDQGPPERV VLLGEFLHPC EDDIVCKCT TDENKVPYFN APVYLENKEQ IGKVDEIFGQ LRDFYFSVKL SENMKASSFK KLQKFYIDPY KLLPLQRFLP R PPGEKGPP RGGGRGGRGG GRGGGGRGGG RGGGFRGGRG GGGGGFRGGR GGGFRGRGH UniProtKB: H/ACA ribonucleoprotein complex subunit 1 |
-Macromolecule #4: H/ACA ribonucleoprotein complex subunit 2
| Macromolecule | Name: H/ACA ribonucleoprotein complex subunit 2 / type: protein_or_peptide / ID: 4 / Number of copies: 2 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 17.22607 KDa |
| Sequence | String: MTKIKADPDG PEAQAEACSG ERTYQELLVN QNPIAQPLAS RRLTRKLYKC IKKAVKQKQI RRGVKEVQKF VNKGEKGIMV LAGDTLPIE VYCHLPVMCE DRNLPYVYIP SKTDLGAAAG SKRPTCVIMV KPHEEYQEAY DECLEEVQSL PLPL UniProtKB: H/ACA ribonucleoprotein complex subunit 2 |
-Macromolecule #5: H/ACA ribonucleoprotein complex subunit 3
| Macromolecule | Name: H/ACA ribonucleoprotein complex subunit 3 / type: protein_or_peptide / ID: 5 / Number of copies: 2 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 7.719989 KDa |
| Sequence | String: MFLQYYLNEQ GDRVYTLKKF DPMGQQTCSA HPARFSPDDK YSRHRITIKK RFKVLMTQQP RPVL UniProtKB: H/ACA ribonucleoprotein complex subunit 3 |
-Macromolecule #6: Telomerase RNA component
| Macromolecule | Name: Telomerase RNA component / type: rna / ID: 6 / Number of copies: 1 |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 145.477797 KDa |
| Sequence | String: GGGUUGCGGA GGGUGGGCCU GGGAGGGGUG GUGGCCAUUU UUUGUCUAAC CCUAACUGAG AAGGGCGUAG GCGCCGUGCU UUUGCUCCC CGCGCGCUGU UUUUCUCGCU GACUUUCAGC GGGCGGAAAA GCCUCGGCCU GCCGCCUUCC ACCGUUCAUU C UAGAGCAA ...String: GGGUUGCGGA GGGUGGGCCU GGGAGGGGUG GUGGCCAUUU UUUGUCUAAC CCUAACUGAG AAGGGCGUAG GCGCCGUGCU UUUGCUCCC CGCGCGCUGU UUUUCUCGCU GACUUUCAGC GGGCGGAAAA GCCUCGGCCU GCCGCCUUCC ACCGUUCAUU C UAGAGCAA ACAAAAAAUG UCAGCUGCUG GCCCGUUCGC CCCUCCCGGG GACCUGCGGC GGGUCGCCUG CCCAGCCCCC GA ACCCCGC CUGGAGGCCG CGGUCGGCCC GGGGCUUCUC CGGAGGCACC CACUGCCACC GCGAAGAGUU GGGCUCUGUC AGC CGCGGG UCUCUCGGGG GCGAGGGCGA GGUUCAGGCC UUUCAGGCCG CAGGAAGAGG AACGGAGCGA GUCCCCGCGC GCGG CGCGA UUCCCUGAGC UGUGGGACGU GCACCCAGGA CUCGGCUCAC ACAUGC GENBANK: GENBANK: U85256.1 |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.9 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 (6k x 4k) / Average electron dose: 62.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
-Image processing
| Startup model | Type of model: INSILICO MODEL |
|---|---|
| Final reconstruction | Resolution.type: BY AUTHOR / Resolution: 3.94 Å / Resolution method: FSC 0.143 CUT-OFF / Number images used: 167923 |
| Initial angle assignment | Type: COMMON LINE |
| Final angle assignment | Type: MAXIMUM LIKELIHOOD |
