National Natural Science Foundation of China (NSFC)
31972287
China
Ministry of Science and Technology (MoST, China)
2017YFD0201400
China
Ministry of Science and Technology (MoST, China)
2017YFD0201403
China
Canadian Institutes of Health Research (CIHR)
PJT-159601
Canada
Citation
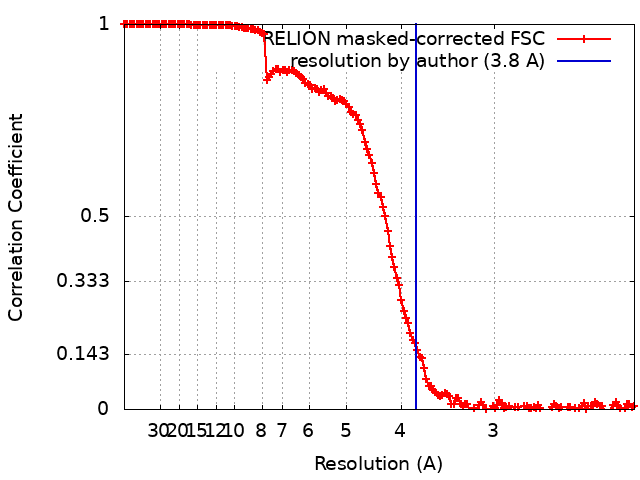
Journal: Nat Chem Biol / Year: 2020 Title: Structural basis for diamide modulation of ryanodine receptor. Authors: Ruifang Ma / Omid Haji-Ghassemi / Dan Ma / Heng Jiang / Lianyun Lin / Li Yao / Arthur Samurkas / Yuxin Li / Yiwen Wang / Peng Cao / Shian Wu / Yan Zhang / Takashi Murayama / Bernard Moussian ...Authors: Ruifang Ma / Omid Haji-Ghassemi / Dan Ma / Heng Jiang / Lianyun Lin / Li Yao / Arthur Samurkas / Yuxin Li / Yiwen Wang / Peng Cao / Shian Wu / Yan Zhang / Takashi Murayama / Bernard Moussian / Filip Van Petegem / Zhiguang Yuchi / Abstract: The diamide insecticide class is one of the top-selling insecticides globally. They are used to control a wide range of pests by targeting their ryanodine receptors (RyRs). Here, we report the ...The diamide insecticide class is one of the top-selling insecticides globally. They are used to control a wide range of pests by targeting their ryanodine receptors (RyRs). Here, we report the highest-resolution cryo-electron microscopy (cryo-EM) structure of RyR1 in the open state, in complex with the anthranilic diamide chlorantraniliprole (CHL). The 3.2-Å local resolution map facilitates unambiguous assignment of the CHL binding site. The molecule induces a conformational change by affecting the S4-S5 linker, triggering channel opening. The binding site is further corroborated by mutagenesis data, which reveal how diamide insecticides are selective to the Lepidoptera group of insects over honeybee or mammalian RyRs. Our data reveal that several pests have developed resistance via two mechanisms, steric hindrance and loss of contact. Our results provide a foundation for the development of highly selective pesticides aimed at overcoming resistance and therapeutic molecules to treat human myopathies.
History
Deposition
Mar 1, 2020
-
Header (metadata) release
Sep 2, 2020
-
Map release
Sep 2, 2020
-
Update
Mar 27, 2024
-
Current status
Mar 27, 2024
Processing site: PDBj / Status: Released
-
Structure visualization
Movie
Surface view with section colored by density value
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Map data
Map data Sample
Sample Keywords
Keywords Function and homology information
Function and homology information
 Homo sapiens (human)
Homo sapiens (human) Authors
Authors China,
China,  Canada, 4 items
Canada, 4 items  Citation
Citation


 Structure visualization
Structure visualization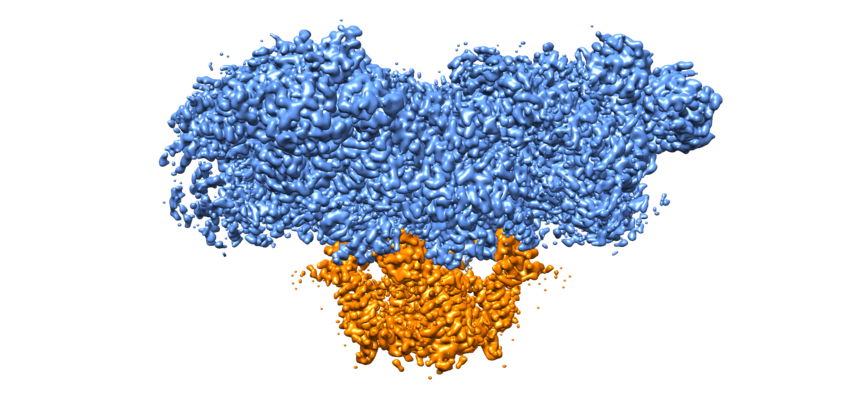
 Downloads & links
Downloads & links emd_30067.png
emd_30067.png http://ftp.pdbj.org/pub/emdb/structures/EMD-30067
http://ftp.pdbj.org/pub/emdb/structures/EMD-30067





























 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)




























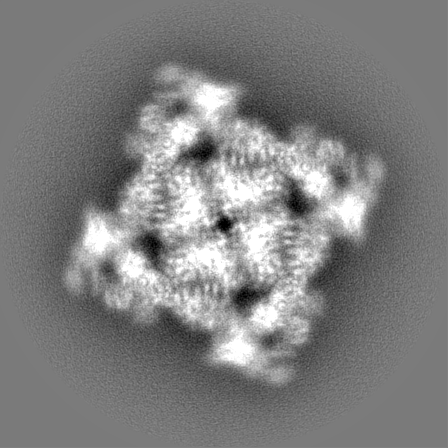

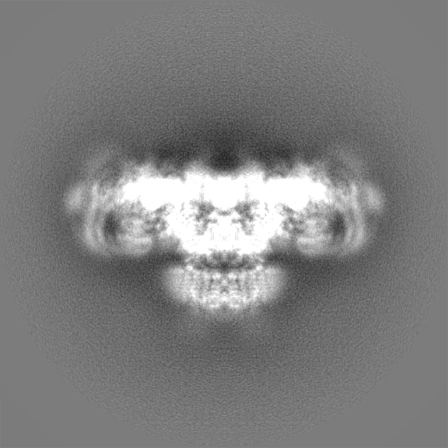
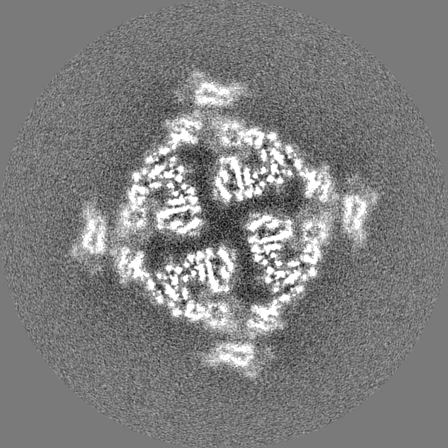
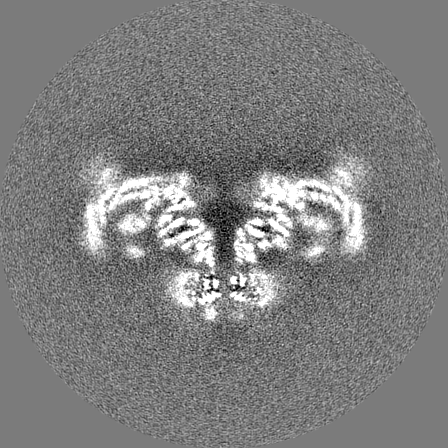
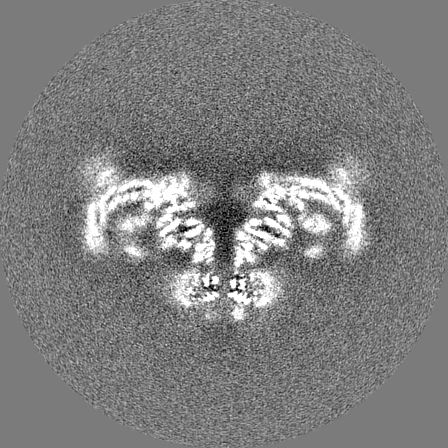
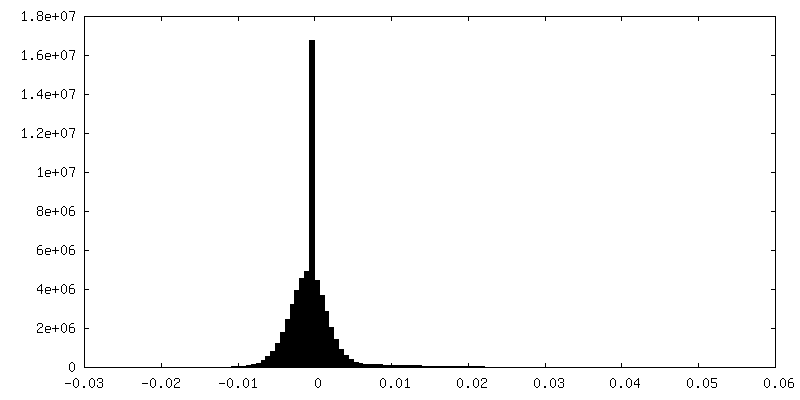
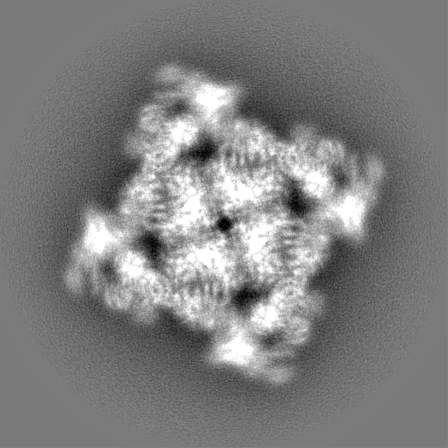
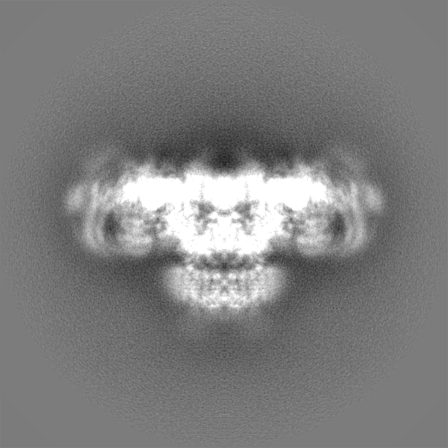
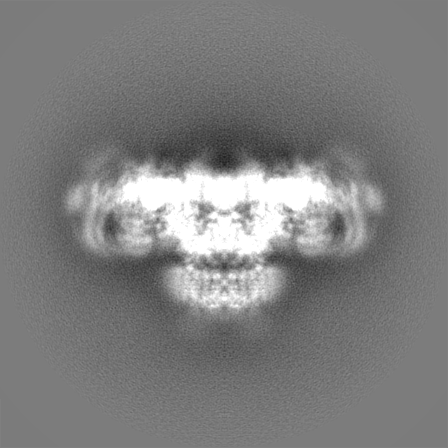
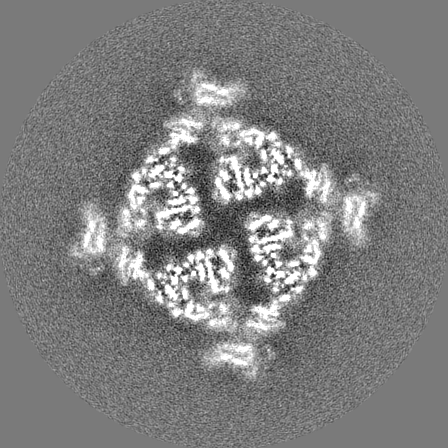
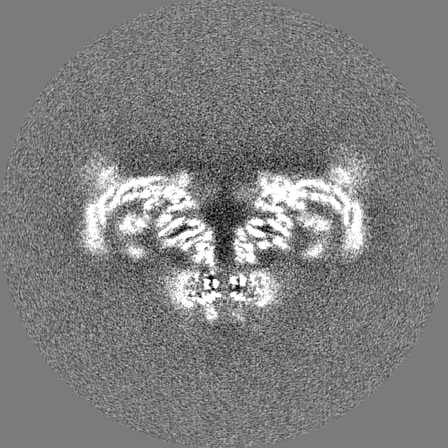
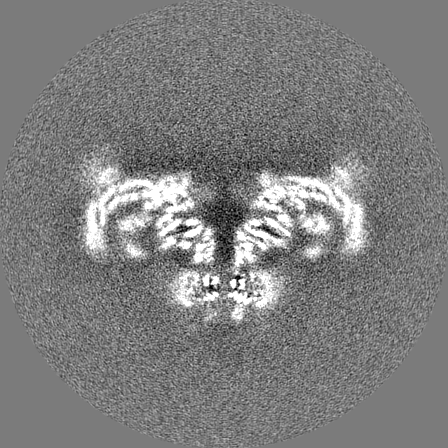
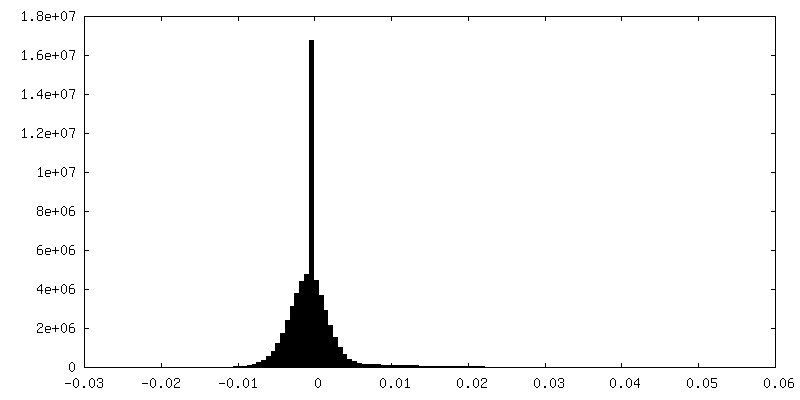
 Sample components
Sample components

 Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN