telomere maintenance via telomere trimming / chromosomal region / telomeric 3' overhang formation / Mre11 complex / establishment of RNA localization to telomere / positive regulation of telomerase catalytic core complex assembly / blastocyst growth / negative regulation of telomere capping / BRCA1-C complex / Sensing of DNA Double Strand Breaks ...telomere maintenance via telomere trimming / chromosomal region / telomeric 3' overhang formation / Mre11 complex / establishment of RNA localization to telomere / positive regulation of telomerase catalytic core complex assembly / blastocyst growth / negative regulation of telomere capping / BRCA1-C complex / Sensing of DNA Double Strand Breaks / cellular response to nitrosative stress / protection from non-homologous end joining at telomere / peptidyl-serine autophosphorylation / establishment of protein-containing complex localization to telomere / meiotic telomere clustering / R-loop processing / t-circle formation / positive regulation of telomere maintenance via telomere lengthening / pre-B cell allelic exclusion / male meiotic nuclear division / phosphorylation-dependent protein binding / histone mRNA catabolic process / DNA-dependent protein kinase activity / extrinsic component of synaptic vesicle membrane / regulation of telomere maintenance via telomerase / histone H2AXS139 kinase activity / female meiotic nuclear division / DNA strand resection involved in replication fork processing / homologous recombination / nuclear inclusion body / DNA double-strand break processing / lipoprotein catabolic process / regulation of autophagosome assembly / cellular response to X-ray / V(D)J recombination / pexophagy / oocyte development / Impaired BRCA2 binding to PALB2 / double-strand break repair via alternative nonhomologous end joining / chromatin-protein adaptor activity / protein localization to site of double-strand break / isotype switching / HDR through MMEJ (alt-NHEJ) / mitotic G2/M transition checkpoint / reciprocal meiotic recombination / DNA repair complex / positive regulation of DNA damage response, signal transduction by p53 class mediator / regulation of DNA-templated DNA replication initiation / Homologous DNA Pairing and Strand Exchange / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / Resolution of D-loop Structures through Holliday Junction Intermediates / 1-phosphatidylinositol-3-kinase activity / HDR through Single Strand Annealing (SSA) / neuromuscular process controlling balance / response to ionizing radiation / positive regulation of double-strand break repair / Impaired BRCA2 binding to RAD51 / mitotic spindle assembly checkpoint signaling / negative regulation of B cell proliferation / TP53 Regulates Transcription of Caspase Activators and Caspases / cellular response to stress / mitotic G2 DNA damage checkpoint signaling / positive regulation of telomere maintenance / TP53 Regulates Transcription of Genes Involved in Cytochrome C Release / telomere maintenance in response to DNA damage / peroxisomal matrix / Presynaptic phase of homologous DNA pairing and strand exchange / replicative senescence / signal transduction in response to DNA damage / protein K63-linked ubiquitination / neuroblast proliferation / Regulation of HSF1-mediated heat shock response / somitogenesis / positive regulation of double-strand break repair via homologous recombination / Pexophagy / ovarian follicle development / regulation of cellular response to heat / cellular response to retinoic acid / negative regulation of TORC1 signaling / positive regulation of telomere maintenance via telomerase / telomere maintenance / positive regulation of cell adhesion / intrinsic apoptotic signaling pathway / DNA damage checkpoint signaling / Stabilization of p53 / thymus development / protein serine/threonine kinase activator activity / regulation of signal transduction by p53 class mediator / replication fork / Autodegradation of the E3 ubiquitin ligase COP1 / meiotic cell cycle / post-embryonic development / determination of adult lifespan / double-strand break repair via homologous recombination / cellular response to reactive oxygen species / TP53 Regulates Transcription of DNA Repair Genes / DNA damage response, signal transduction by p53 class mediator 類似検索 - 分子機能
Nibrin, C-terminal / Nibrin / DNA damage repair protein Nbs1 / DNA damage repair protein Nbs1 / Nibrin, second BRCT domain / Nibrin, second BRCT domain superfamily / Second BRCT domain on Nijmegen syndrome breakage protein / Nibrin-related / Telomere-length maintenance and DNA damage repair / Serine/threonine-protein kinase ATM, plant ...Nibrin, C-terminal / Nibrin / DNA damage repair protein Nbs1 / DNA damage repair protein Nbs1 / Nibrin, second BRCT domain / Nibrin, second BRCT domain superfamily / Second BRCT domain on Nijmegen syndrome breakage protein / Nibrin-related / Telomere-length maintenance and DNA damage repair / Serine/threonine-protein kinase ATM, plant / ATM, catalytic domain / Telomere-length maintenance and DNA damage repair / Telomere-length maintenance and DNA damage repair / Forkhead associated domain / Forkhead-associated (FHA) domain profile. / FATC domain / FHA domain / PIK-related kinase, FAT / FAT domain / Forkhead-associated (FHA) domain / FATC / FATC domain / PIK-related kinase / FAT domain profile. / FATC domain profile. / SMAD/FHA domain superfamily / BRCA1 C Terminus (BRCT) domain / Phosphatidylinositol 3- and 4-kinases signature 1. / Phosphatidylinositol 3/4-kinase, conserved site / Phosphatidylinositol 3- and 4-kinases signature 2. / Phosphatidylinositol 3-/4-kinase, catalytic domain superfamily / Phosphoinositide 3-kinase, catalytic domain / Phosphatidylinositol 3- and 4-kinase / Phosphatidylinositol 3- and 4-kinases catalytic domain profile. / Phosphatidylinositol 3-/4-kinase, catalytic domain / BRCT domain / BRCT domain superfamily / Armadillo-type fold / Protein kinase-like domain superfamily 類似検索 - ドメイン・相同性
National Institutes of Health/National Cancer Institute (NIH/NCI)
5F32CA247320
米国
National Institutes of Health/Eunice Kennedy Shriver National Institute of Child Health & Human Development (NIH/NICHD)
CA008748
米国
引用
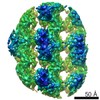
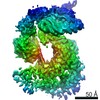
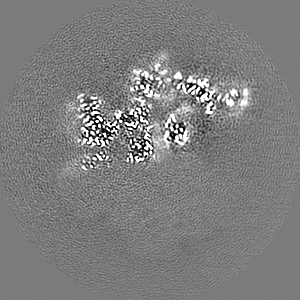
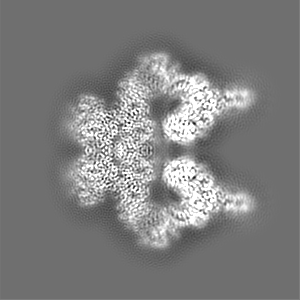
ジャーナル: Elife / 年: 2022 タイトル: Structure of the human ATM kinase and mechanism of Nbs1 binding. 著者: Christopher Warren / Nikola P Pavletich / 要旨: DNA double-strand breaks (DSBs) can lead to mutations, chromosomal rearrangements, genome instability, and cancer. Central to the sensing of DSBs is the ATM (Ataxia-telangiectasia mutated) kinase, ...DNA double-strand breaks (DSBs) can lead to mutations, chromosomal rearrangements, genome instability, and cancer. Central to the sensing of DSBs is the ATM (Ataxia-telangiectasia mutated) kinase, which belongs to the phosphatidylinositol 3-kinase-related protein kinase (PIKK) family. In response to DSBs, ATM is activated by the MRN (Mre11-Rad50-Nbs1) protein complex through a poorly understood process that also requires double-stranded DNA. Previous studies indicate that the FxF/Y motif of Nbs1 directly binds to ATM, and is required to retain active ATM at sites of DNA damage. Here, we report the 2.5 Å resolution cryo-EM structures of human ATM and its complex with the Nbs1 FxF/Y motif. In keeping with previous structures of ATM and its yeast homolog Tel1, the dimeric human ATM kinase adopts a symmetric, butterfly-shaped structure. The conformation of the ATM kinase domain is most similar to the inactive states of other PIKKs, suggesting that activation may involve an analogous realigning of the N and C lobes along with relieving the blockage of the substrate-binding site. We also show that the Nbs1 FxF/Y motif binds to a conserved hydrophobic cleft within the Spiral domain of ATM, suggesting an allosteric mechanism of activation. We evaluate the importance of these structural findings with mutagenesis and biochemical assays.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 米国, 2件
米国, 2件  引用
引用 構造の表示
構造の表示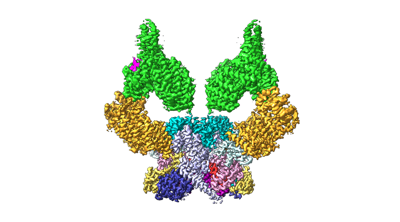
 ダウンロードとリンク
ダウンロードとリンク emd_25141.png
emd_25141.png http://ftp.pdbj.org/pub/emdb/structures/EMD-25141
http://ftp.pdbj.org/pub/emdb/structures/EMD-25141























 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)



























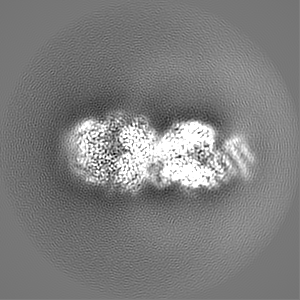
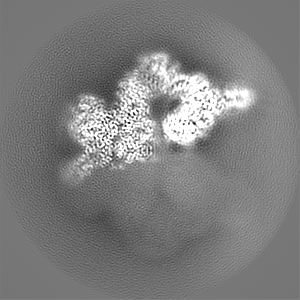
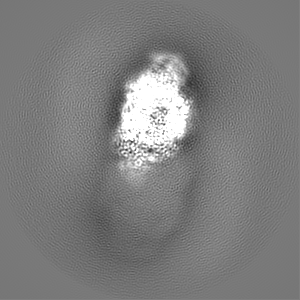
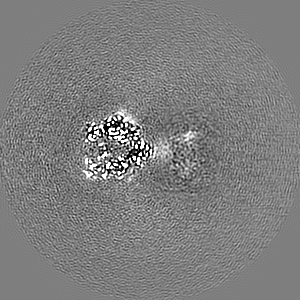



 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN