negative regulation of DNA-templated DNA replication / regulation of cellular response to hypoxia / negative regulation of receptor signaling pathway via JAK-STAT / negative regulation of beige fat cell differentiation / RHOBTB3 ATPase cycle / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling ...negative regulation of DNA-templated DNA replication / regulation of cellular response to hypoxia / negative regulation of receptor signaling pathway via JAK-STAT / negative regulation of beige fat cell differentiation / RHOBTB3 ATPase cycle / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / target-directed miRNA degradation / cellular response to chemical stress / elongin complex / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / Replication of the SARS-CoV-1 genome / protein neddylation / transcription elongation factor activity / NEDD8 ligase activity / negative regulation of response to oxidative stress / protein K27-linked ubiquitination / VCB complex / Cul5-RING ubiquitin ligase complex / ribosome-associated ubiquitin-dependent protein catabolic process / ubiquitin-ubiquitin ligase activity / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / SUMOylation of ubiquitinylation proteins / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / negative regulation of mitophagy / Prolactin receptor signaling / histone H4K8ac reader activity / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / negative regulation of transcription elongation by RNA polymerase II / Cul4B-RING E3 ubiquitin ligase complex / RNA polymerase II C-terminal domain binding / histone H3K27ac reader activity / negative regulation of DNA damage checkpoint / ubiquitin ligase complex scaffold activity / P-TEFb complex binding / histone H3K9ac reader activity / histone H4 reader activity / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / histone H4K5ac reader activity / histone H4K12ac reader activity / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / host-mediated suppression of viral transcription / histone H4K16ac reader activity / positive regulation of G2/M transition of mitotic cell cycle / protein monoubiquitination / cullin family protein binding / negative regulation of signal transduction / positive regulation of T-helper 17 cell lineage commitment / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / ubiquitin-like ligase-substrate adaptor activity / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / signal transduction in response to DNA damage / negative regulation of TORC1 signaling / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / negative regulation of insulin receptor signaling pathway / Ribosome Quality Control (RQC) complex extracts and degrades nascent peptide / RNA polymerase II CTD heptapeptide repeat kinase activity / RNA Polymerase II Pre-transcription Events / regulation of cellular response to insulin stimulus / positive regulation of TORC1 signaling / rescue of stalled cytosolic ribosome / intrinsic apoptotic signaling pathway / transcription-coupled nucleotide-excision repair / post-translational protein modification / negative regulation of autophagy / transcription corepressor binding / negative regulation of canonical NF-kappaB signal transduction / cellular response to amino acid stimulus / site of DNA damage / condensed nuclear chromosome / Regulation of BACH1 activity / protein serine/threonine kinase binding / T cell activation / positive regulation of cell differentiation / TP53 Regulates Transcription of DNA Repair Genes / transcription coregulator activity / G1/S transition of mitotic cell cycle / positive regulation of transcription elongation by RNA polymerase II / negative regulation of canonical Wnt signaling pathway / cell morphogenesis / transcription initiation at RNA polymerase II promoter / Degradation of DVL / Degradation of CRY and PER proteins / transcription elongation by RNA polymerase II / Degradation of GLI1 by the proteasome Similarity search - Function
European Union, United Kingdom, Switzerland, 4 items
Organization
Grant number
Country
European Research Council (ERC)
European Union
Wellcome Trust
United Kingdom
Medical Research Council (MRC, United Kingdom)
United Kingdom
Innovative Medicines Initiative
Switzerland
Citation
Journal: Sci Adv / Year: 2024 Title: Mechanism of degrader-targeted protein ubiquitinability. Authors: Charlotte Crowe / Mark A Nakasone / Sarah Chandler / Conner Craigon / Gajanan Sathe / Michael H Tatham / Nikolai Makukhin / Ronald T Hay / Alessio Ciulli / Abstract: Small-molecule degraders of disease-driving proteins offer a clinically proven modality with enhanced therapeutic efficacy and potential to tackle previously undrugged targets. Stable and long-lived ...Small-molecule degraders of disease-driving proteins offer a clinically proven modality with enhanced therapeutic efficacy and potential to tackle previously undrugged targets. Stable and long-lived degrader-mediated ternary complexes drive fast and profound target degradation; however, the mechanisms by which they affect target ubiquitination remain elusive. Here, we show cryo-EM structures of the VHL Cullin 2 RING E3 ligase with the degrader MZ1 directing target protein Brd4 toward UBE2R1-ubiquitin, and Lys at optimal positioning for nucleophilic attack. In vitro ubiquitination and mass spectrometry illuminate a patch of favorably ubiquitinable lysines on one face of Brd4, with cellular degradation and ubiquitinomics confirming the importance of Lys and nearby Lys/Lys, identifying the "ubiquitination zone." Our results demonstrate the proficiency of MZ1 in positioning the substrate for catalysis, the favorability of Brd4 for ubiquitination by UBE2R1, and the flexibility of CRL2 for capturing suboptimal lysines. We propose a model for ubiquitinability of degrader-recruited targets, providing a mechanistic blueprint for further rational drug design.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information
 Map data
Map data Sample
Sample Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) Authors
Authors United Kingdom,
United Kingdom,  Switzerland, 4 items
Switzerland, 4 items  Citation
Citation Structure visualization
Structure visualization
 Downloads & links
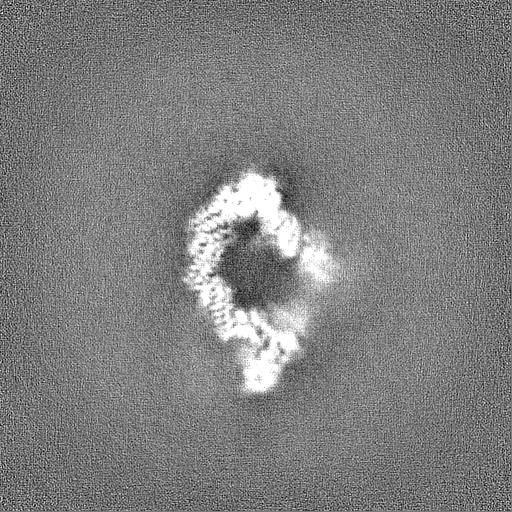
Downloads & links emd_19569.png
emd_19569.png http://ftp.pdbj.org/pub/emdb/structures/EMD-19569
http://ftp.pdbj.org/pub/emdb/structures/EMD-19569

















 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)
































 Sample components
Sample components Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN