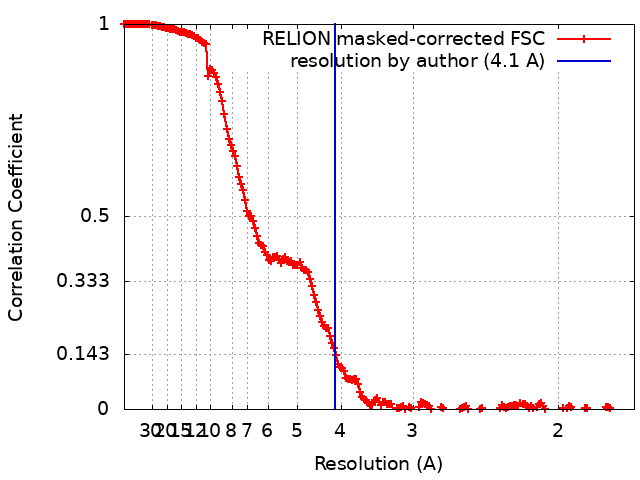
- EMDB-18216: Structure of CUL9-RBX1 ubiquitin E3 ligase complex in unneddylate... -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: EMDB / ID: EMD-18216
タイトル
Structure of CUL9-RBX1 ubiquitin E3 ligase complex in unneddylated and neddylated conformation - focused cullin dimer
マップデータ
DeepEMhancer map
試料
複合体: Structure of CUL9-RBX1 ubiquitin E3 ligase complex in unneddylated and neddylated conformation - focused cullin dimer
タンパク質・ペプチド: Cullin-9
タンパク質・ペプチド: E3 ubiquitin-protein ligase RBX1
タンパク質・ペプチド: NEDD8
タンパク質・ペプチド: Unknown
リガンド: ZINC ION
キーワード
Cullin-RING RBR E3 Ligase / LIGASE
機能・相同性
機能・相同性情報
negative regulation of beige fat cell differentiation / cellular response to camptothecin / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance ...negative regulation of beige fat cell differentiation / cellular response to camptothecin / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / protein neddylation / NEDD8 ligase activity / protein K27-linked ubiquitination / negative regulation of response to oxidative stress / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-ubiquitin ligase activity / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / regulation of mitotic nuclear division / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / Prolactin receptor signaling / negative regulation of mitophagy / Cul4B-RING E3 ubiquitin ligase complex / TGF-beta receptor signaling activates SMADs / regulation of proteolysis / regulation of postsynapse assembly / cullin family protein binding / anatomical structure morphogenesis / protein monoubiquitination / site of DNA damage / signal transduction in response to DNA damage / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / transcription-coupled nucleotide-excision repair / negative regulation of insulin receptor signaling pathway / regulation of cellular response to insulin stimulus / positive regulation of TORC1 signaling / post-translational protein modification / T cell activation / protein modification process / Regulation of BACH1 activity / negative regulation of canonical NF-kappaB signal transduction / cellular response to amino acid stimulus / Degradation of DVL / Degradation of CRY and PER proteins / negative regulation of canonical Wnt signaling pathway / Iron uptake and transport / Degradation of GLI1 by the proteasome / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Recognition of DNA damage by PCNA-containing replication complex / Negative regulation of NOTCH4 signaling / RING-type E3 ubiquitin transferase / Hedgehog 'on' state / Vif-mediated degradation of APOBEC3G / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / NOTCH1 Intracellular Domain Regulates Transcription / Degradation of beta-catenin by the destruction complex / Evasion by RSV of host interferon responses / modification-dependent protein catabolic process / DNA Damage Recognition in GG-NER / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / microtubule cytoskeleton organization / Constitutive Signaling by NOTCH1 PEST Domain Mutants / Constitutive Signaling by NOTCH1 HD+PEST Domain Mutants / Dual Incision in GG-NER / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / Formation of TC-NER Pre-Incision Complex / Regulation of expression of SLITs and ROBOs / Formation of Incision Complex in GG-NER / Interleukin-1 signaling / protein tag activity / Orc1 removal from chromatin / protein polyubiquitination / Dual incision in TC-NER / Regulation of RAS by GAPs / Gap-filling DNA repair synthesis and ligation in TC-NER / ubiquitin-protein transferase activity / Regulation of RUNX2 expression and activity / positive regulation of protein catabolic process / cellular response to UV / intracellular protein localization / ubiquitin protein ligase activity / UCH proteinases / KEAP1-NFE2L2 pathway / MAPK cascade / Cargo recognition for clathrin-mediated endocytosis / Antigen processing: Ubiquitination & Proteasome degradation / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / Neddylation / cellular response to oxidative stress / spermatogenesis 類似検索 - 分子機能
ジャーナル: Nat Struct Mol Biol / 年: 2024 タイトル: Noncanonical assembly, neddylation and chimeric cullin-RING/RBR ubiquitylation by the 1.8 MDa CUL9 E3 ligase complex. 著者: Daniel Horn-Ghetko / Linus V M Hopf / Ishita Tripathi-Giesgen / Jiale Du / Sebastian Kostrhon / D Tung Vu / Viola Beier / Barbara Steigenberger / J Rajan Prabu / Luca Stier / Elias M Bruss / ...著者: Daniel Horn-Ghetko / Linus V M Hopf / Ishita Tripathi-Giesgen / Jiale Du / Sebastian Kostrhon / D Tung Vu / Viola Beier / Barbara Steigenberger / J Rajan Prabu / Luca Stier / Elias M Bruss / Matthias Mann / Yue Xiong / Brenda A Schulman / 要旨: Ubiquitin ligation is typically executed by hallmark E3 catalytic domains. Two such domains, 'cullin-RING' and 'RBR', are individually found in several hundred human E3 ligases, and collaborate with ...Ubiquitin ligation is typically executed by hallmark E3 catalytic domains. Two such domains, 'cullin-RING' and 'RBR', are individually found in several hundred human E3 ligases, and collaborate with E2 enzymes to catalyze ubiquitylation. However, the vertebrate-specific CUL9 complex with RBX1 (also called ROC1), of interest due to its tumor suppressive interaction with TP53, uniquely encompasses both cullin-RING and RBR domains. Here, cryo-EM, biochemistry and cellular assays elucidate a 1.8-MDa hexameric human CUL9-RBX1 assembly. Within one dimeric subcomplex, an E2-bound RBR domain is activated by neddylation of its own cullin domain and positioning from the adjacent CUL9-RBX1 in trans. Our data show CUL9 as unique among RBX1-bound cullins in dependence on the metazoan-specific UBE2F neddylation enzyme, while the RBR domain protects it from deneddylation. Substrates are recruited to various upstream domains, while ubiquitylation relies on both CUL9's neddylated cullin and RBR domains achieving self-assembled and chimeric cullin-RING/RBR E3 ligase activity.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 ドイツ, 2件
ドイツ, 2件  引用
引用
 構造の表示
構造の表示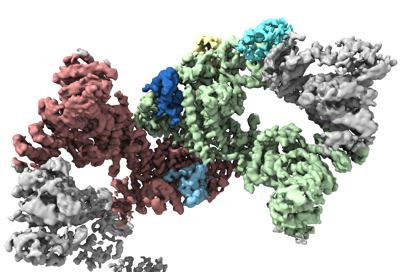
 ダウンロードとリンク
ダウンロードとリンク emd_18216.png
emd_18216.png http://ftp.pdbj.org/pub/emdb/structures/EMD-18216
http://ftp.pdbj.org/pub/emdb/structures/EMD-18216
















 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)



































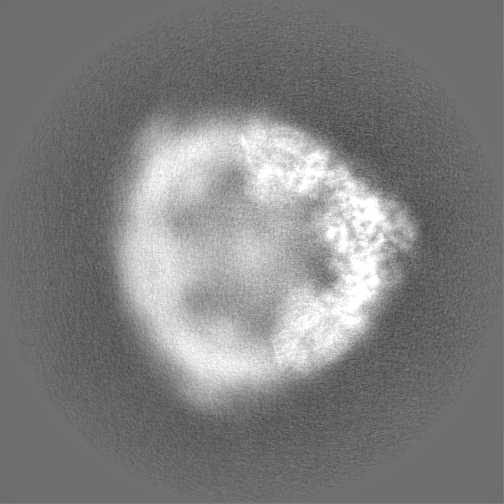
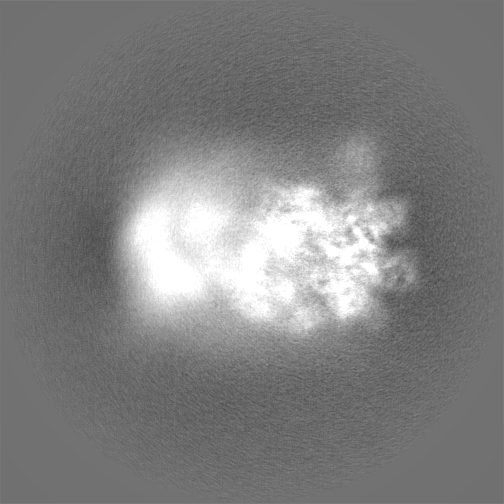
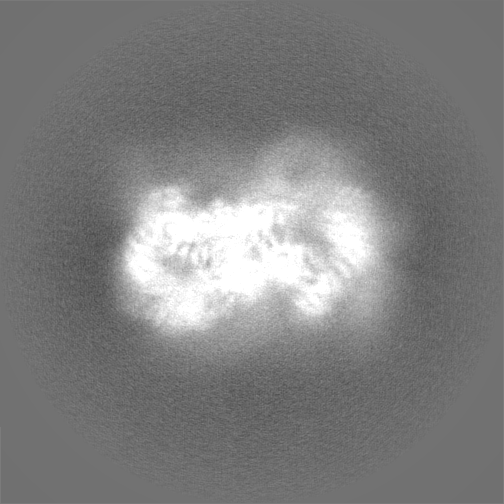

 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN