ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
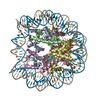
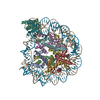
| タイトル | Nucleosome Bound human SIRT6 (composite structure) | |||||||||
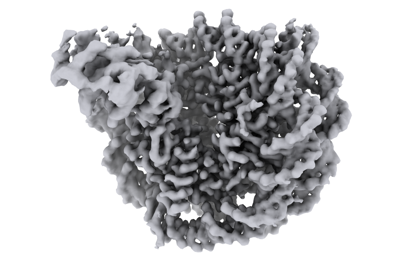
 マップデータ マップデータ | Composite map | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Transferase / Deacetylase / Histone H3 deacetylation / GENE REGULATION | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報histone H3K56 deacetylase activity, NAD-dependent / histone H3K18 deacetylase activity, NAD-dependent / ketone biosynthetic process / histone H3K9 deacetylase activity, hydrolytic mechanism / histone H3K9 deacetylase activity, NAD-dependent / protein delipidation / NAD+-protein-lysine ADP-ribosyltransferase activity / regulation of lipid catabolic process / chromosome, subtelomeric region / positive regulation of protein localization to chromatin ...histone H3K56 deacetylase activity, NAD-dependent / histone H3K18 deacetylase activity, NAD-dependent / ketone biosynthetic process / histone H3K9 deacetylase activity, hydrolytic mechanism / histone H3K9 deacetylase activity, NAD-dependent / protein delipidation / NAD+-protein-lysine ADP-ribosyltransferase activity / regulation of lipid catabolic process / chromosome, subtelomeric region / positive regulation of protein localization to chromatin / NAD+-protein-arginine ADP-ribosyltransferase activity / DNA damage sensor activity / NAD-dependent protein demyristoylase activity / NAD-dependent protein depalmitoylase activity / positive regulation of stem cell differentiation / negative regulation of D-glucose import / positive regulation of chondrocyte proliferation / transposable element silencing / NAD-dependent protein lysine deacetylase activity / cardiac muscle cell differentiation / protein acetyllysine N-acetyltransferase / pericentric heterochromatin formation / protein deacetylation / histone deacetylase activity, NAD-dependent / protein localization to site of double-strand break / positive regulation of blood vessel branching / negative regulation of glycolytic process / TORC2 complex binding / negative regulation of protein localization to chromatin / positive regulation of vascular endothelial cell proliferation / histone deacetylase regulator activity / negative regulation of protein import into nucleus / regulation of double-strand break repair via homologous recombination / regulation of protein secretion / positive regulation of double-strand break repair / DNA repair-dependent chromatin remodeling / positive regulation of stem cell proliferation / lncRNA binding / negative regulation of gene expression, epigenetic / NAD+-protein mono-ADP-ribosyltransferase activity / positive regulation of stem cell population maintenance / positive regulation of telomere maintenance / regulation of lipid metabolic process / site of DNA damage / negative regulation of transcription elongation by RNA polymerase II / negative regulation of cellular senescence / NAD+ poly-ADP-ribosyltransferase activity / 転移酵素; グリコシル基を移すもの; 五炭糖残基を移すもの / NAD+ binding / negative regulation of gluconeogenesis / positive regulation of fat cell differentiation / subtelomeric heterochromatin formation / pericentric heterochromatin / response to UV / regulation of protein localization to plasma membrane / nucleosome binding / enzyme regulator activity / 転移酵素; アシル基を移すもの; アミノアシル基以外のアシル基を移すもの / nucleotidyltransferase activity / positive regulation of protein export from nucleus / determination of adult lifespan / circadian regulation of gene expression / base-excision repair / regulation of circadian rhythm / protein destabilization / positive regulation of insulin secretion / chromatin DNA binding / Pre-NOTCH Transcription and Translation / positive regulation of fibroblast proliferation / protein import into nucleus / structural constituent of chromatin / transcription corepressor activity / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / nucleosome / heterochromatin formation / glucose homeostasis / double-strand break repair / nucleosome assembly / site of double-strand break / positive regulation of cold-induced thermogenesis / Processing of DNA double-strand break ends / damaged DNA binding / chromatin remodeling / protein heterodimerization activity / negative regulation of cell population proliferation / intracellular membrane-bounded organelle / chromatin binding / chromatin / endoplasmic reticulum / negative regulation of transcription by RNA polymerase II / protein homodimerization activity / DNA binding / zinc ion binding / nucleoplasm / nucleus 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.94 Å | |||||||||
 データ登録者 データ登録者 | Smirnova E / Bignon E / Schultz P / Papai G / Ben-Shem A | |||||||||
| 資金援助 |  フランス, 1件 フランス, 1件
| |||||||||
 引用 引用 | ジャーナル: Elife / 年: 2024 タイトル: Binding to nucleosome poises human SIRT6 for histone H3 deacetylation. 著者: Ekaterina Smirnova / Emmanuelle Bignon / Patrick Schultz / Gabor Papai / Adam Ben Shem / 要旨: Sirtuin 6 (SIRT6) is an NAD-dependent histone H3 deacetylase that is prominently found associated with chromatin, attenuates transcriptionally active promoters and regulates DNA repair, metabolic ...Sirtuin 6 (SIRT6) is an NAD-dependent histone H3 deacetylase that is prominently found associated with chromatin, attenuates transcriptionally active promoters and regulates DNA repair, metabolic homeostasis and lifespan. Unlike other sirtuins, it has low affinity to free histone tails but demonstrates strong binding to nucleosomes. It is poorly understood how SIRT6 docking on nucleosomes stimulates its histone deacetylation activity. Here, we present the structure of human SIRT6 bound to a nucleosome determined by cryogenic electron microscopy. The zinc finger domain of SIRT6 associates tightly with the acidic patch of the nucleosome through multiple arginine anchors. The Rossmann fold domain binds to the terminus of the looser DNA half of the nucleosome, detaching two turns of the DNA from the histone octamer and placing the NAD binding pocket close to the DNA exit site. This domain shows flexibility with respect to the fixed zinc finger and moves with, but also relative to, the unwrapped DNA terminus. We apply molecular dynamics simulations of the histone tails in the nucleosome to show that in this mode of interaction, the active site of SIRT6 is perfectly poised to catalyze deacetylation of the H3 histone tail and that the partial unwrapping of the DNA allows even lysines close to the H3 core to reach the enzyme. #1: ジャーナル: Elife / 年: 2023タイトル: Binding to nucleosome poises SIRT6 for histone H3 de-acetylation 著者: Smirnova E / Bignon E / Schultz P / Papai G / Ben-Shem A | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_16845.map.gz | 38.5 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-16845-v30.xmlemd-16845.xml | 23.7 KB 23.7 KB | 表示 表示 | EMDBヘッダ |
| 画像 | 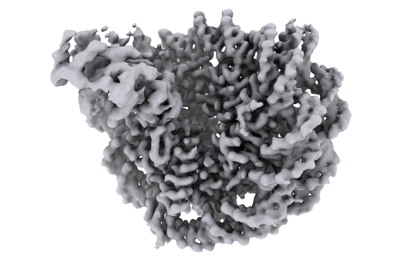 emd_16845.png emd_16845.png | 97.2 KB | ||
| Filedesc metadata | emd-16845.cif.gz | 7 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-16845ftp://ftp.pdbj.org/pub/emdb/structures/EMD-16845 http://ftp.pdbj.org/pub/emdb/structures/EMD-16845ftp://ftp.pdbj.org/pub/emdb/structures/EMD-16845 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_16845_validation.pdf.gz | 398.9 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_16845_full_validation.pdf.gz | 398.5 KB | 表示 | |
| XML形式データ | emd_16845_validation.xml.gz | 5.6 KB | 表示 | |
| CIF形式データ | emd_16845_validation.cif.gz | 6.4 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-16845ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-16845 | HTTPS FTP |
-関連構造データ




-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |




-マップ
| ファイル | ダウンロード / ファイル: emd_16845.map.gz / 形式: CCP4 / 大きさ: 42.9 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Composite map | ||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 0.916 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)


















-添付データ
- 試料の構成要素
試料の構成要素
+全体 : Human Sirtuin 6 in complex with the nucleosome
+超分子 #1: Human Sirtuin 6 in complex with the nucleosome
+超分子 #2: SIRT6
+分子 #1: Histone H3.2

+分子 #2: Histone H4
+分子 #3: Histone H2A type 1
+分子 #4: Histone H2B
+分子 #7: NAD-dependent protein deacylase sirtuin-6
+分子 #5: DNA (145-MER)
+分子 #6: DNA (145-MER)
+分子 #8: ZINC ION
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.5 |
|---|---|
| グリッド | モデル: Quantifoil R3.5/1 / 材質: COPPER/RHODIUM / メッシュ: 300 / 支持フィルム - 材質: CARBON / 支持フィルム - トポロジー: CONTINUOUS / 支持フィルム - Film thickness: 2 |
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法 #1
電子顕微鏡法 #1
| Microscopy ID | 1 |
|---|---|
| 顕微鏡 | TFS KRIOS |
| 特殊光学系 | エネルギーフィルター - 名称: TFS Selectris X / エネルギーフィルター - スリット幅: 10 eV |
| 撮影 | Image recording ID: 1 フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) 平均電子線量: 50.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 50.0 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 2.6 µm / 最小 デフォーカス(公称値): 1.2 µm / 倍率(公称値): 270000 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-電子顕微鏡法 #1~
| Microscopy ID | 1 |
|---|---|
| 顕微鏡 | TFS KRIOS |
| 特殊光学系 | エネルギーフィルター - 名称: GIF Quantum LS / エネルギーフィルター - スリット幅: 20 eV |
| 撮影 | Image recording ID: 2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 平均電子線量: 55.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源: FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 50.0 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 0.01 mm / 最大 デフォーカス(公称値): 2.6 µm / 最小 デフォーカス(公称値): 1.2 µm / 倍率(公称値): 180000 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 | モデル: Titan Krios / 画像提供: FEI Company |