 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
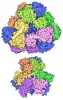
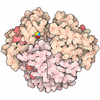
Yorodumi- EMDB-14119: Structure of the human MHC I peptide-loading complex editing module -
+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of the human MHC I peptide-loading complex editing module | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | antigen processing / peptide proofreading / chaperones / MHC I / IMMUNE SYSTEM | |||||||||
| Function / homology |  Function and homology information Function and homology informationMHC class Ib protein complex assembly / peptide antigen stabilization / Tapasin-ERp57 complex / response to biphenyl / Calnexin/calreticulin cycle / MHC class I protein complex binding / cytolytic granule / antigen processing and presentation of exogenous peptide antigen via MHC class I, TAP-dependent / nuclear receptor-mediated glucocorticoid signaling pathway / protein disulfide-isomerase ...MHC class Ib protein complex assembly / peptide antigen stabilization / Tapasin-ERp57 complex / response to biphenyl / Calnexin/calreticulin cycle / MHC class I protein complex binding / cytolytic granule / antigen processing and presentation of exogenous peptide antigen via MHC class I, TAP-dependent / nuclear receptor-mediated glucocorticoid signaling pathway / protein disulfide-isomerase / Assembly of Viral Components at the Budding Site / positive regulation of dendritic cell chemotaxis / ATF6 (ATF6-alpha) activates chaperone genes / negative regulation of trophoblast cell migration / cortical granule / regulation of meiotic nuclear division / cellular response to electrical stimulus / : / negative regulation of retinoic acid receptor signaling pathway / complement component C1q complex binding / endoplasmic reticulum quality control compartment / protein folding in endoplasmic reticulum / sarcoplasmic reticulum lumen / response to peptide / negative regulation of intracellular steroid hormone receptor signaling pathway / disulfide oxidoreductase activity / TAP2 binding / TAP1 binding / regulation of protein complex stability / nuclear export signal receptor activity / C-type glycerophospholipase activity / retrograde vesicle-mediated transport, Golgi to endoplasmic reticulum / cardiac muscle cell differentiation / cortical actin cytoskeleton organization / cellular response to interleukin-7 / positive regulation of memory T cell activation / T cell mediated cytotoxicity directed against tumor cell target / positive regulation of extrinsic apoptotic signaling pathway / response to glycoside / positive regulation of CD8-positive, alpha-beta T cell activation / CD8-positive, alpha-beta T cell activation / positive regulation of CD8-positive, alpha-beta T cell proliferation / Scavenging by Class A Receptors / Scavenging by Class F Receptors / cellular response to lithium ion / nuclear androgen receptor binding / negative regulation of neuron differentiation / antigen processing and presentation of endogenous peptide antigen via MHC class I via ER pathway, TAP-dependent / TAP complex binding / antigen processing and presentation of exogenous peptide antigen via MHC class I / protein disulfide isomerase activity / Golgi medial cisterna / response to testosterone / lncRNA binding / CD8 receptor binding / smooth endoplasmic reticulum / hormone binding / protection from natural killer cell mediated cytotoxicity / protein-disulfide reductase activity / beta-2-microglobulin binding / endoplasmic reticulum exit site / TAP binding / MHC class I protein binding / molecular sequestering activity / detection of bacterium / antigen processing and presentation of endogenous peptide antigen via MHC class Ib / antigen processing and presentation of endogenous peptide antigen via MHC class I via ER pathway, TAP-independent / protein localization to nucleus / extrinsic apoptotic signaling pathway / T cell receptor binding / phagocytic vesicle / endocytic vesicle lumen / peptide binding / ERAD pathway / positive regulation of cell cycle / positive regulation of substrate adhesion-dependent cell spreading / endoplasmic reticulum-Golgi intermediate compartment membrane / protein export from nucleus / protein folding chaperone / positive regulation of endothelial cell migration / positive regulation of phagocytosis / early endosome lumen / acrosomal vesicle / Nef mediated downregulation of MHC class I complex cell surface expression / DAP12 interactions / response to endoplasmic reticulum stress / Endosomal/Vacuolar pathway / T cell mediated cytotoxicity / Antigen Presentation: Folding, assembly and peptide loading of class I MHC / lumenal side of endoplasmic reticulum membrane / regulation of iron ion transport / cellular response to iron(III) ion / negative regulation of iron ion transport / negative regulation of forebrain neuron differentiation / antigen processing and presentation of exogenous protein antigen via MHC class Ib, TAP-dependent / peptide antigen assembly with MHC class I protein complex / regulation of erythrocyte differentiation / ER to Golgi transport vesicle membrane / response to molecule of bacterial origin / HFE-transferrin receptor complex Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.73 Å | |||||||||
 Authors Authors | Domnick A / Susac L | |||||||||
| Funding support | European Union, 2 items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2022 Title: Molecular basis of MHC I quality control in the peptide loading complex. Authors: Alexander Domnick / Christian Winter / Lukas Sušac / Leon Hennecke / Mario Hensen / Nicole Zitzmann / Simon Trowitzsch / Christoph Thomas / Robert Tampé /   Abstract: Major histocompatibility complex class I (MHC I) molecules are central to adaptive immunity. Their assembly, epitope selection, and antigen presentation are controlled by the MHC I glycan through a ...Major histocompatibility complex class I (MHC I) molecules are central to adaptive immunity. Their assembly, epitope selection, and antigen presentation are controlled by the MHC I glycan through a sophisticated network of chaperones and modifying enzymes. However, the mechanistic integration of the corresponding processes remains poorly understood. Here, we determine the multi-chaperone-client interaction network of the peptide loading complex (PLC) and report the PLC editing module structure by cryogenic electron microscopy at 3.7 Å resolution. Combined with epitope-proofreading studies of the PLC in near-native lipid environment, these data show that peptide-receptive MHC I molecules are stabilized by multivalent chaperone interactions including the calreticulin-engulfed mono-glucosylated MHC I glycan, which only becomes accessible for processing by α-glucosidase II upon loading of optimal epitopes. Our work reveals allosteric coupling between peptide-MHC I assembly and glycan processing. This inter-process communication defines the onset of an adaptive immune response and provides a prototypical example of the tightly coordinated events in endoplasmic reticulum quality control. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_14119.map.gz | 166.7 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-14119-v30.xmlemd-14119.xml | 14.1 KB 14.1 KB | Display Display | EMDB header |
| Images |  emd_14119.png emd_14119.png | 70.6 KB | ||
| Filedesc metadata | emd-14119.cif.gz | 6.4 KB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-14119ftp://ftp.pdbj.org/pub/emdb/structures/EMD-14119 http://ftp.pdbj.org/pub/emdb/structures/EMD-14119ftp://ftp.pdbj.org/pub/emdb/structures/EMD-14119 | HTTPS FTP |
-Related structure data
| Related structure data |  7qpdMC M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |
-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |







-Map
| File | Download / File: emd_14119.map.gz / Format: CCP4 / Size: 178 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.05 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
- Sample components
Sample components
-Entire : editing module of peptide-loading complex
| Entire | Name: editing module of peptide-loading complex |
|---|---|
| Components |
|
-Supramolecule #1: editing module of peptide-loading complex
| Supramolecule | Name: editing module of peptide-loading complex / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
-Macromolecule #1: Beta-2-microglobulin
| Macromolecule | Name: Beta-2-microglobulin / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 11.74816 KDa |
| Sequence | String: IQRTPKIQVY SRHPAENGKS NFLNCYVSGF HPSDIEVDLL KNGERIEKVE HSDLSFSKDW SFYLLYYTEF TPTEKDEYAC RVNHVTLSQ PKIVKWDRDM UniProtKB: Beta-2-microglobulin |
-Macromolecule #2: Tapasin
| Macromolecule | Name: Tapasin / type: protein_or_peptide / ID: 2 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 45.761184 KDa |
| Sequence | String: GPAVIECWFV EDASGKGLAK RPGALLLRQG PGEPPPRPDL DPELYLSVHD PAGALQAAFR RYPRGAPAPH CEMSRFVPLP ASAKWASGL TPAQNCPRAL DGAWLMVSIS SPVLSLSSLL RPQPEPQQEP VLITMATVVL TVLTHTPAPR VRLGQDALLD L SFAYMPPT ...String: GPAVIECWFV EDASGKGLAK RPGALLLRQG PGEPPPRPDL DPELYLSVHD PAGALQAAFR RYPRGAPAPH CEMSRFVPLP ASAKWASGL TPAQNCPRAL DGAWLMVSIS SPVLSLSSLL RPQPEPQQEP VLITMATVVL TVLTHTPAPR VRLGQDALLD L SFAYMPPT SEAASSLAPG PPPFGLEWRR QHLGKGHLLL AATPGLNGQM PAAQEGAVAF AAWDDDEPWG PWTGNGTFWL PR VQPFQEG TYLATIHLPY LQGQVTLELA VYKPPKVSLM PATLARAAPG EAPPELLCLV SHFYPSGGLE VEWELRGGPG GRS QKAEGQ RWLSALRHHS DGSVSLSGHL QPPPVTTEQH GARYACRIHH PSLPASGRSA EVTLEVAGLS GPSLEDSVGL FLSA FLLLG LFKALGWAAV YLSTCKDSKK KAE UniProtKB: Tapasin |
-Macromolecule #3: Protein disulfide-isomerase A3
| Macromolecule | Name: Protein disulfide-isomerase A3 / type: protein_or_peptide / ID: 3 / Number of copies: 1 / Enantiomer: LEVO / EC number: protein disulfide-isomerase |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 54.341102 KDa |
| Sequence | String: SDVLELTDDN FESRISDTGS AGLMLVEFFA PWCGHCKRLA PEYEAAATRL KGIVPLAKVD CTANTNTCNK YGVSGYPTLK IFRDGEEAG AYDGPRTADG IVSHLKKQAG PASVPLRTEE EFKKFISDKD ASIVGFFDDS FSEAHSEFLK AASNLRDNYR F AHTNVESL ...String: SDVLELTDDN FESRISDTGS AGLMLVEFFA PWCGHCKRLA PEYEAAATRL KGIVPLAKVD CTANTNTCNK YGVSGYPTLK IFRDGEEAG AYDGPRTADG IVSHLKKQAG PASVPLRTEE EFKKFISDKD ASIVGFFDDS FSEAHSEFLK AASNLRDNYR F AHTNVESL VNEYDDNGEG IILFRPSHLT NKFEDKTVAY TEQKMTSGKI KKFIQENIFG ICPHMTEDNK DLIQGKDLLI AY YDVDYEK NAKGSNYWRN RVMMVAKKFL DAGHKLNFAV ASRKTFSHEL SDFGLESTAG EIPVVAIRTA KGEKFVMQEE FSR DGKALE RFLQDYFDGN LKRYLKSEPI PESNDGPVKV VVAENFDEIV NNENKDVLIE FYAPWCGHCK NLEPKYKELG EKLS KDPNI VIAKMDATAN DVPSPYEVRG FPTIYFSPAN KKLNPKKYEG GRELSDFISY LQREATNPPV IQEEKPKKKK KAQED L UniProtKB: Protein disulfide-isomerase A3 |
-Macromolecule #4: HLA class I histocompatibility antigen, A-3 alpha chain
| Macromolecule | Name: HLA class I histocompatibility antigen, A-3 alpha chain type: protein_or_peptide / ID: 4 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 38.363535 KDa |
| Sequence | String: GSHSMRYFFT SVSRPGRGEP RFIAVGYVDD TQFVRFDSDA ASQRMEPRAP WIEQEGPEYW DQETRNVKAQ SQTDRVDLGT LRGYYNQSE AGSHTIQIMY GCDVGSDGRF LRGYRQDAYD GKDYIALNED LRSWTAADMA AQITKRKWEA AHEAEQLRAY L DGTCVEWL ...String: GSHSMRYFFT SVSRPGRGEP RFIAVGYVDD TQFVRFDSDA ASQRMEPRAP WIEQEGPEYW DQETRNVKAQ SQTDRVDLGT LRGYYNQSE AGSHTIQIMY GCDVGSDGRF LRGYRQDAYD GKDYIALNED LRSWTAADMA AQITKRKWEA AHEAEQLRAY L DGTCVEWL RRYLENGKET LQRTDPPKTH MTHHPISDHE ATLRCWALGF YPAEITLTWQ RDGEDQTQDT ELVETRPAGD GT FQKWAAV VVPSGEEQRY TCHVQHEGLP KPLTLRWELS SQPTIPIVGI IAGLVLLGAV ITGAVVAAVM WRRKSSDRKG GSY TQAASS DSAQGSDVSL TACKV UniProtKB: HLA class I histocompatibility antigen, A alpha chain |
-Macromolecule #5: Calreticulin
| Macromolecule | Name: Calreticulin / type: protein_or_peptide / ID: 5 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 46.523211 KDa |
| Sequence | String: EPAVYFKEQF LDGDGWTSRW IESKHKSDFG KFVLSSGKFY GDEEKDKGLQ TSQDARFYAL SASFEPFSNK GQTLVVQFTV KHEQNIDCG GGYVKLFPNS LDQTDMHGDS EYNIMFGPDI CGPGTKKVHV IFNYKGKNVL INKDIRCKDD EFTHLYTLIV R PDNTYEVK ...String: EPAVYFKEQF LDGDGWTSRW IESKHKSDFG KFVLSSGKFY GDEEKDKGLQ TSQDARFYAL SASFEPFSNK GQTLVVQFTV KHEQNIDCG GGYVKLFPNS LDQTDMHGDS EYNIMFGPDI CGPGTKKVHV IFNYKGKNVL INKDIRCKDD EFTHLYTLIV R PDNTYEVK IDNSQVESGS LEDDWDFLPP KKIKDPDASK PEDWDERAKI DDPTDSKPED WDKPEHIPDP DAKKPEDWDE EM DGEWEPP VIQNPEYKGE WKPRQIDNPD YKGTWIHPEI DNPEYSPDPS IYAYDNFGVL GLDLWQVKSG TIFDNFLITN DEA YAEEFG NETWGVTKAA EKQMKDKQDE EQRLKEEEED KKRKEEEEAE DKEDDEDKDE DEEDEEDKEE DEEEDVPGQA KDEL UniProtKB: Calreticulin |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.4 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Average electron dose: 68.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 2.5 µm / Nominal defocus min: 1.0 µm |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
-Image processing
| Startup model | Type of model: NONE |
|---|---|
| Final reconstruction | Resolution.type: BY AUTHOR / Resolution: 3.73 Å / Resolution method: FSC 0.143 CUT-OFF / Number images used: 97952 |
| Initial angle assignment | Type: MAXIMUM LIKELIHOOD |
| Final angle assignment | Type: MAXIMUM LIKELIHOOD |
