 Movie
Movie Controller
Controller
[English] 日本語
 Yorodumi
Yorodumi- SASDE96: Aldehyde dehydrogenase 12 from Zea mays Extrapolated to Infinite ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: SASBDB / ID: SASDE96 |
|---|---|
 Sample Sample | Aldehyde dehydrogenase 12 from Zea mays Extrapolated to Infinite Dilution
|
| Function / homology |  Function and homology information Function and homology informationL-glutamate gamma-semialdehyde dehydrogenase / L-glutamate gamma-semialdehyde dehydrogenase activity / aldehyde dehydrogenase (NAD+) activity / nucleotide binding Similarity search - Function |
| Biological species |  |
 Citation Citation | Journal: J Mol Biol / Year: 2019 Title: Structural and Biochemical Characterization of Aldehyde Dehydrogenase 12, the Last Enzyme of Proline Catabolism in Plants. Authors: David A Korasick / Radka Končitíková / Martina Kopečná / Eva Hájková / Armelle Vigouroux / Solange Moréra / Donald F Becker / Marek Šebela / John J Tanner / David Kopečný /    Abstract: Heterokonts, Alveolata protists, green algae from Charophyta and Chlorophyta divisions, and all Embryophyta plants possess an aldehyde dehydrogenase (ALDH) gene named ALDH12. Here, we provide a ...Heterokonts, Alveolata protists, green algae from Charophyta and Chlorophyta divisions, and all Embryophyta plants possess an aldehyde dehydrogenase (ALDH) gene named ALDH12. Here, we provide a biochemical characterization of two ALDH12 family members from the lower plant Physcomitrella patens and higher plant Zea mays. We show that ALDH12 encodes an NAD-dependent glutamate γ-semialdehyde dehydrogenase (GSALDH), which irreversibly converts glutamate γ-semialdehyde (GSAL), a mitochondrial intermediate of the proline and arginine catabolism, to glutamate. Sedimentation equilibrium and small-angle X-ray scattering analyses reveal that in solution both plant GSALDHs exist as equilibrium between a domain-swapped dimer and the dimer-of-dimers tetramer. Plant GSALDHs share very low-sequence identity with bacterial, fungal, and animal GSALDHs (classified as ALDH4), which are the closest related ALDH superfamily members. Nevertheless, the crystal structure of ZmALDH12 at 2.2-Å resolution shows that nearly all key residues involved in the recognition of GSAL are identical to those in ALDH4, indicating a close functional relationship with ALDH4. Phylogenetic analysis suggests that the transition from ALDH4 to ALDH12 occurred during the evolution of the endosymbiotic plant ancestor, prior to the evolution of green algae and land plants. Finally, ALDH12 expression in maize and moss is downregulated in response to salt and drought stresses, possibly to maintain proline levels. Taken together, these results provide molecular insight into the biological roles of the plant ALDH12 family. |
 Contact author Contact author |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Data source
| SASBDB page |  SASDE96 SASDE96 |
|---|
-Related structure data




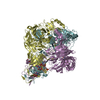
-External links
| Related items in Molecule of the Month |
|---|
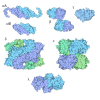

-Models
| Model #2495 |   Type: mix / Chi-square value: 0.577963245374368 / P-value: 0.000012  Search similar-shape structures of this assembly by Omokage search (details) Search similar-shape structures of this assembly by Omokage search (details) |
|---|---|
| Model #2494 |   Type: mix / Chi-square value: 57.3783426075885 Search similar-shape structures of this assembly by Omokage search (details) |
-Sample
| Sample | Name: Aldehyde dehydrogenase 12 from Zea mays Extrapolated to Infinite Dilution |
|---|---|
| Buffer | Name: 50 mM Tris-HCl, 50 mM NaCl, 0.5 mM TCEP, and 5% (v/v) glycerol pH: 7.8 |
| Entity #1332 | Type: protein / Description: Aldehyde dehydrogenase 12 / Formula weight: 60.397 / Num. of mol.: 4 / Source: Zea mays / References: UniProt: A0A2H4PMI3 Sequence: MGSSHHHHHH SQDPAPFACV SRWLHTPSFA TVSPQEVSGS SPAEVQNFVQ GSWTASANWN WIVDPLNGDK FIKVAEVQGT EIKSFMESLS KCPKHGLHNP LKAPERYLMY GDISAKAAHM LGQPTVLDFF AKLIQRVSPK SYQQALAEVQ VSQKFLENFC GDQVRFLARS ...Sequence: MGSSHHHHHH SQDPAPFACV SRWLHTPSFA TVSPQEVSGS SPAEVQNFVQ GSWTASANWN WIVDPLNGDK FIKVAEVQGT EIKSFMESLS KCPKHGLHNP LKAPERYLMY GDISAKAAHM LGQPTVLDFF AKLIQRVSPK SYQQALAEVQ VSQKFLENFC GDQVRFLARS FAVPGNHLGQ RSNGYRWPYG PVAIITPFNF PLEIPLLQLM GALYMGNKPV LKVDSKVSIV MEQMIRLLHD CGLPAEDMDF INSDGAVMNK LLLEANPKMT LFTGSSRVAE KLAADLKGRV KLEDAGFDWK ILGPDVQEVD YVAWVCDQDA YACSGQKCSA QSVLFMHKNW SSSGLLEKMK KLSERRKLED LTIGPVLTVT TEAMIEHMNN LLKIRGSKVL FGGEPLANHS IPKIYGAMKP TAVFVPLEEI LKSGNFELVT KEIFGPFQVV TEYSEDQLEL VLEACERMNA HLTAAIVSND PLFLQDVLGR SVNGTTYAGI RARTTGAPQN HWFGPAGDPR GAGIGTPEAI KLVWSCHREI IYDVGPVPES WALPSAT |
-Experimental information
| Beam | Instrument name: Advanced Light Source (ALS) 12.3.1 (SIBYLS) City: Berkeley, CA / 国: USA / Type of source: X-ray synchrotron / Wavelength: 0.127 Å / Dist. spec. to detc.: 1.5 mm | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Detector | Name: Pilatus3 X 2M / Pixsize x: 172 mm | ||||||||||||||||||||||||||||||
| Scan |  Measurement date: Dec 6, 2016 / Storage temperature: 4 °C / Cell temperature: 10 °C / Exposure time: 0.3 sec. / Number of frames: 33 / Unit: 1/A /
| ||||||||||||||||||||||||||||||
| Distance distribution function P(R) | 
| ||||||||||||||||||||||||||||||
| Result |  Comments: This is the background subtracted experimental SAXS data extrapolated to infinite dilution. The original experiment consisted of static SAXS samples collected at 3 protein concentrations in ...Comments: This is the background subtracted experimental SAXS data extrapolated to infinite dilution. The original experiment consisted of static SAXS samples collected at 3 protein concentrations in the range of 0.9 - 2.6 mg/mL. The data were extrapolated to infinite dilution to account for sample aggregation in the two higher concentration samples. The corresponding model fits of the tetramer (top) and dimer (bottom) demonstrate that the dimer does not fit the SAXS data.
|
