- PDB-9bfn: Cryo-EM co-structure of AcrB with the CU232 efflux pump inhibitor -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: PDB / ID: 9bfn
タイトル
Cryo-EM co-structure of AcrB with the CU232 efflux pump inhibitor
要素
Multidrug efflux pump subunit AcrB
キーワード
TRANSLOCASE / AcrB Multidrug Efflux Pump
機能・相同性
機能・相同性情報
alkane transmembrane transporter activity / alkane transport / enterobactin transport / enterobactin transmembrane transporter activity / xenobiotic detoxification by transmembrane export across the cell outer membrane / periplasmic side of plasma membrane / efflux pump complex / Iron assimilation using enterobactin / bile acid transmembrane transporter activity / Antimicrobial resistance ...alkane transmembrane transporter activity / alkane transport / enterobactin transport / enterobactin transmembrane transporter activity / xenobiotic detoxification by transmembrane export across the cell outer membrane / periplasmic side of plasma membrane / efflux pump complex / Iron assimilation using enterobactin / bile acid transmembrane transporter activity / Antimicrobial resistance / bile acid and bile salt transport / Secretion of toxins / efflux transmembrane transporter activity / xenobiotic transmembrane transporter activity / fatty acid transport / xenobiotic transport / response to toxic substance / response to xenobiotic stimulus / response to antibiotic / membrane / identical protein binding / plasma membrane 類似検索 - 分子機能
National Institutes of Health/National Institute Of Allergy and Infectious Diseases (NIH/NIAID)
米国
引用
ジャーナル: mBio / 年: 2023 タイトル: Bacterial efflux pump modulators prevent bacterial growth in macrophages and under broth conditions that mimic the host environment. 著者: Samual C Allgood / Chih-Chia Su / Amy L Crooks / Christian T Meyer / Bojun Zhou / Meredith D Betterton / Michael R Barbachyn / Edward W Yu / Corrella S Detweiler / 要旨: Bacterial efflux pumps are critical for resistance to antibiotics and for virulence. We previously identified small molecules that inhibit efflux pumps (efflux pump modulators, EPMs) and prevent ...Bacterial efflux pumps are critical for resistance to antibiotics and for virulence. We previously identified small molecules that inhibit efflux pumps (efflux pump modulators, EPMs) and prevent pathogen replication in host cells. Here, we used medicinal chemistry to increase the activity of the EPMs against pathogens in cells into the nanomolar range. We show by cryo-electron microscopy that these EPMs bind an efflux pump subunit. In broth culture, the EPMs increase the potency (activity), but not the efficacy (maximum effect), of antibiotics. We also found that bacterial exposure to the EPMs appear to enable the accumulation of a toxic metabolite that would otherwise be exported by efflux pumps. Thus, inhibitors of bacterial efflux pumps could interfere with infection not only by potentiating antibiotics, but also by allowing toxic waste products to accumulate within bacteria, providing an explanation for why efflux pumps are needed for virulence in the absence of antibiotics.
履歴
登録
2024年4月18日
登録サイト: RCSB / 処理サイト: RCSB
改定 1.0
2024年5月8日
Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: EM metadata / Data content type: EM metadata / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: Additional map / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: FSC / Data content type: FSC / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: Half map / Part number: 1 / Data content type: Half map / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: Half map / Part number: 2 / Data content type: Half map / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年5月8日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
Data content type: EM metadata / Data content type: EM metadata / EM metadata / Group: Data processing / Experimental summary / Data content type: EM metadata / EM metadata / カテゴリ: em_admin / em_software / Data content type: EM metadata / EM metadata / Item: _em_admin.last_update / _em_software.name
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 米国, 1件
米国, 1件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード






 PDBj
PDBj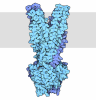
 集合体
集合体

 分子量: 748.065 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C41H82NO8P / コメント: リン脂質*YM
分子量: 748.065 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C41H82NO8P / コメント: リン脂質*YM 分子量: 18.015 Da / 分子数: 5 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 5 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 Homo sapiens (ヒト)
Homo sapiens (ヒト) 電子顕微鏡撮影
電子顕微鏡撮影
 FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN
FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN 解析
解析