 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8p03 | ||||||
|---|---|---|---|---|---|---|---|
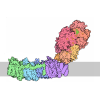
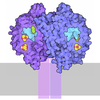
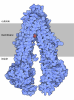
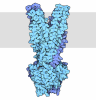
| タイトル | 48S late-stage initiation complex with m6A mRNA | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | TRANSLATION / translation initiation / ribosome / m6A / methylated mRNA | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of macromolecule metabolic process / Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / Major pathway of rRNA processing in the nucleolus and cytosol / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / SRP-dependent cotranslational protein targeting to membrane / Formation of a pool of free 40S subunits / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) ...regulation of macromolecule metabolic process / Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / Major pathway of rRNA processing in the nucleolus and cytosol / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / SRP-dependent cotranslational protein targeting to membrane / Formation of a pool of free 40S subunits / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / eukaryotic translation initiation factor 2 complex / eukaryotic 48S preinitiation complex / cellular response to chemical stress / laminin receptor activity / 90S preribosome / ubiquitin ligase inhibitor activity / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / positive regulation of signal transduction by p53 class mediator / regulation of translational fidelity / phagocytic cup / translation regulator activity / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / rough endoplasmic reticulum / ribosomal small subunit export from nucleus / laminin binding / translation initiation factor activity / gastrulation / MDM2/MDM4 family protein binding / cytosolic ribosome / class I DNA-(apurinic or apyrimidinic site) endonuclease activity / response to endoplasmic reticulum stress / DNA-(apurinic or apyrimidinic site) lyase / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / positive regulation of apoptotic signaling pathway / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / small-subunit processome / spindle / cytoplasmic stress granule / rRNA processing / regulation of translation / rhythmic process / positive regulation of canonical Wnt signaling pathway / ribosomal small subunit assembly / virus receptor activity / ribosome binding / ribosomal small subunit biogenesis / small ribosomal subunit / small ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / perikaryon / cell differentiation / cytoplasmic translation / tRNA binding / mitochondrial inner membrane / postsynaptic density / rRNA binding / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / cell division / DNA repair / mRNA binding / apoptotic process / centrosome / synapse / dendrite / nucleolus / perinuclear region of cytoplasm / Golgi apparatus / ATP hydrolysis activity / DNA binding / RNA binding / zinc ion binding / ATP binding / membrane / nucleus / plasma membrane / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.04 Å | ||||||
 データ登録者 データ登録者 | Guca, E. / Lima, L.H.F. / Boissier, F. / Hashem, Y. | ||||||
| 資金援助 | European Union, 1件
| ||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2024 タイトル: N-methyladenosine in 5' UTR does not promote translation initiation. 著者: Ewelina Guca / Rodrigo Alarcon / Michael Z Palo / Leonardo Santos / Santiago Alonso-Gil / Marcos Davyt / Leonardo H F de Lima / Fanny Boissier / Sarada Das / Bojan Zagrovic / Joseph D Puglisi ...著者: Ewelina Guca / Rodrigo Alarcon / Michael Z Palo / Leonardo Santos / Santiago Alonso-Gil / Marcos Davyt / Leonardo H F de Lima / Fanny Boissier / Sarada Das / Bojan Zagrovic / Joseph D Puglisi / Yaser Hashem / Zoya Ignatova /      要旨: The most abundant N-methyladenosine (mA) modification on mRNAs is installed non-stoichiometrically across transcripts, with 5' untranslated regions (5' UTRs) being the least conductive. 5' UTRs are ...The most abundant N-methyladenosine (mA) modification on mRNAs is installed non-stoichiometrically across transcripts, with 5' untranslated regions (5' UTRs) being the least conductive. 5' UTRs are essential for translation initiation, yet the molecular mechanisms orchestrated by mA remain poorly understood. Here, we combined structural, biochemical, and single-molecule approaches and show that at the most common position, a single mA does not affect translation yields, the kinetics of translation initiation complex assembly, or start codon recognition both under permissive growth and following exposure to oxidative stress. Cryoelectron microscopy (cryo-EM) structures of the late preinitiation complex reveal that mA purine ring established stacking interactions with an arginine side chain of the initiation factor eIF2α, although with only a marginal energy contribution, as estimated computationally. These findings provide molecular insights into mA interactions with the initiation complex and suggest that the subtle stabilization is unlikely to affect the translation dynamics under homeostatic conditions or stress. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8p03.cif.gz | 1.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8p03.ent.gz | 1.5 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8p03.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/p0/8p03ftp://data.pdbj.org/pub/pdb/validation_reports/p0/8p03 | HTTPS FTP |
|---|
-関連構造データ



-リンク
 PDBj
PDBj




































- 集合体
集合体
| 登録構造単位 | 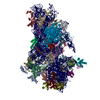
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 3種, 3分子 123
| #1: RNA鎖 | 分子量: 24376.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: RNA鎖 | 分子量: 601015.688 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: RNA鎖 | 分子量: 2882.836 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) |
-Eukaryotic translation initiation factor ... , 2種, 2分子 Aj
| #4: タンパク質 | 分子量: 32792.867 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #37: タンパク質 | 分子量: 12788.764 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+40S ribosomal protein ... , 25種, 25分子 CEGIKLMNOQRSTUVWXZabcdein
-Ribosomal protein ... , 8種, 8分子 DFHJPYfg
| #6: タンパク質 | 分子量: 24944.408 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #8: タンパク質 | 分子量: 25158.535 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) 参照: UniProt: G1TNM3, DNA-(apurinic or apyrimidinic site) lyase |
| #10: タンパク質 | 分子量: 21525.941 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 21716.387 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 17128.191 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #27: タンパク質 | 分子量: 14865.555 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #34: タンパク質 | 分子量: 8358.903 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #35: タンパク質 | 分子量: 34669.113 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 / タンパク質・ペプチド / 非ポリマー , 3種, 180分子 kl
| #38: タンパク質 | 分子量: 66986.812 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #39: タンパク質・ペプチド | 分子量: 3473.451 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #41: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 178 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 178 / 由来タイプ: 合成 / 式: Mg |
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: 48S late-stage initiation complex with m6A-modified mRNA タイプ: RIBOSOME / Entity ID: #1-#40 / 由来: NATURAL |
|---|---|
| 由来(天然) | 生物種: |
| 緩衝液 | pH: 7.4 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Talos Arctica / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TECNAI ARCTICA |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: OTHER / 最大 デフォーカス(公称値): 2300 nm / 最小 デフォーカス(公称値): 300 nm |
| 撮影 | 電子線照射量: 45 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION |
|---|---|
| 3次元再構成 | 解像度: 3.04 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 103050 / 対称性のタイプ: POINT |
