ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8gae | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
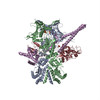
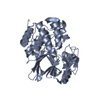
| タイトル | Hsp90 provides platform for CRaf dephosphorylation by PP5 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 キーワード キーワード | CHAPERONE / kinase / phosphatase / complex | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of type II interferon-mediated signaling pathway / peptidyl-serine dephosphorylation / positive regulation of nuclear receptor-mediated glucocorticoid signaling pathway / response to arachidonate / HSP90-CDC37 chaperone complex / death-inducing signaling complex assembly / negative regulation of proteasomal protein catabolic process / peptidyl-threonine dephosphorylation / Aryl hydrocarbon receptor signalling / aryl hydrocarbon receptor complex ...regulation of type II interferon-mediated signaling pathway / peptidyl-serine dephosphorylation / positive regulation of nuclear receptor-mediated glucocorticoid signaling pathway / response to arachidonate / HSP90-CDC37 chaperone complex / death-inducing signaling complex assembly / negative regulation of proteasomal protein catabolic process / peptidyl-threonine dephosphorylation / Aryl hydrocarbon receptor signalling / aryl hydrocarbon receptor complex / intermediate filament cytoskeleton organization / histone methyltransferase binding / dynein axonemal particle / receptor ligand inhibitor activity / regulation of Rho protein signal transduction / proximal dendrite / positive regulation of type 2 mitophagy / type B pancreatic cell proliferation / SHOC2 M1731 mutant abolishes MRAS complex function / Gain-of-function MRAS complexes activate RAF signaling / regulation of cyclin-dependent protein serine/threonine kinase activity / ATP-dependent protein binding / positive regulation of protein localization to cell surface / Rap1 signalling / protein kinase regulator activity / insulin secretion involved in cellular response to glucose stimulus / mitogen-activated protein kinase kinase kinase binding / protein folding chaperone complex / Negative feedback regulation of MAPK pathway / response to morphine / IFNG signaling activates MAPKs / post-transcriptional regulation of gene expression / GP1b-IX-V activation signalling / protein-serine/threonine phosphatase / Respiratory syncytial virus genome replication / telomerase holoenzyme complex assembly / ERBB2-ERBB3 signaling pathway / positive regulation of transforming growth factor beta receptor signaling pathway / Uptake and function of diphtheria toxin / neurotrophin TRK receptor signaling pathway / Drug-mediated inhibition of ERBB2 signaling / Resistance of ERBB2 KD mutants to trastuzumab / Resistance of ERBB2 KD mutants to sapitinib / Resistance of ERBB2 KD mutants to tesevatinib / Resistance of ERBB2 KD mutants to neratinib / Resistance of ERBB2 KD mutants to osimertinib / Resistance of ERBB2 KD mutants to afatinib / Resistance of ERBB2 KD mutants to AEE788 / Resistance of ERBB2 KD mutants to lapatinib / Drug resistance in ERBB2 TMD/JMD mutants / regulation of type I interferon-mediated signaling pathway / pseudopodium / dendritic growth cone / TPR domain binding / face development / cellular response to cadmium ion / protein serine/threonine phosphatase activity / Assembly and release of respiratory syncytial virus (RSV) virions / phosphatase activity / regulation of cell differentiation / thyroid gland development / phosphoprotein phosphatase activity / The NLRP3 inflammasome / protein phosphatase activator activity / Sema3A PAK dependent Axon repulsion / protein serine/threonine kinase inhibitor activity / extrinsic apoptotic signaling pathway via death domain receptors / G-protein alpha-subunit binding / somatic stem cell population maintenance / regulation of protein ubiquitination / positive regulation of peptidyl-serine phosphorylation / HSF1-dependent transactivation / MAP kinase kinase kinase activity / response to unfolded protein / HSF1 activation / type II interferon-mediated signaling pathway / telomere maintenance via telomerase / Attenuation phase / chaperone-mediated protein complex assembly / protein targeting / axonal growth cone / negative regulation of protein-containing complex assembly / Schwann cell development / RHOBTB2 GTPase cycle / Purinergic signaling in leishmaniasis infection / negative regulation of extrinsic apoptotic signaling pathway via death domain receptors / protein dephosphorylation / negative regulation of MAPK cascade / supramolecular fiber organization / : / DNA polymerase binding / heat shock protein binding / response to muscle stretch / Signaling by ERBB2 / Hsp70 protein binding / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / myelination / protein folding chaperone / peptide binding / CD209 (DC-SIGN) signaling 類似検索 - 分子機能 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.3 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Jaime-Garza, M. / Nowotny, C.A. / Coutandin, D. / Wang, F. / Tabios, M. / Agard, D.A. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 資金援助 |  米国, 4件 米国, 4件
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2023 タイトル: Hsp90 provides a platform for kinase dephosphorylation by PP5. 著者: Maru Jaime-Garza / Carlos A Nowotny / Daniel Coutandin / Feng Wang / Mariano Tabios / David A Agard / 要旨: The Hsp90 molecular chaperone collaborates with the phosphorylated Cdc37 cochaperone for the folding and activation of its many client kinases. As with many kinases, the Hsp90 client kinase CRaf is ...The Hsp90 molecular chaperone collaborates with the phosphorylated Cdc37 cochaperone for the folding and activation of its many client kinases. As with many kinases, the Hsp90 client kinase CRaf is activated by phosphorylation at specific regulatory sites. The cochaperone phosphatase PP5 dephosphorylates CRaf and Cdc37 in an Hsp90-dependent manner. Although dephosphorylating Cdc37 has been proposed as a mechanism for releasing Hsp90-bound kinases, here we show that Hsp90 bound kinases sterically inhibit Cdc37 dephosphorylation indicating kinase release must occur before Cdc37 dephosphorylation. Our cryo-EM structure of PP5 in complex with Hsp90:Cdc37:CRaf reveals how Hsp90 both activates PP5 and scaffolds its association with the bound CRaf to dephosphorylate phosphorylation sites neighboring the kinase domain. Thus, we directly show how Hsp90's role in maintaining protein homeostasis goes beyond folding and activation to include post translationally modifying its client kinases. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8gae.cif.gz | 748.5 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8gae.ent.gz | 615 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8gae.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8gae_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8gae_full_validation.pdf.gz | 1.3 MB | 表示 | |
| XML形式データ | 8gae_validation.xml.gz | 71.5 KB | 表示 | |
| CIF形式データ | 8gae_validation.cif.gz | 109.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ga/8gaeftp://data.pdbj.org/pub/pdb/validation_reports/ga/8gae | HTTPS FTP |
-関連構造データ







-リンク
 PDBj
PDBj
















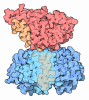

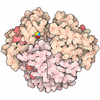
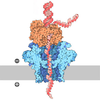




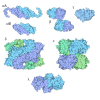

- 集合体
集合体
| 登録構造単位 | 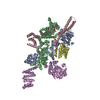
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 5分子 ABCDE
| #1: タンパク質 | 分子量: 83595.484 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: Sequence starts with HRV 3C cleavage site, followed by human Hsp90B sequence. 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HSP90AB1, HSP90B, HSPC2, HSPCB / 発現宿主:  #2: タンパク質 | | 分子量: 45352.223 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CDC37, CDC37A / 発現宿主: #3: タンパク質 | | 分子量: 23841.605 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RAF1, RAF / 発現宿主: 参照: UniProt: P04049, non-specific serine/threonine protein kinase #4: タンパク質 | | 分子量: 57184.730 Da / 分子数: 1 / Mutation: H304A / 由来タイプ: 組換発現 詳細: Sequence starts with HRV 3C cleavage site, continued by human PP5 sequence. 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PPP5C, PPP5 / 発現宿主:  参照: UniProt: P53041, protein-serine/threonine phosphatase |
|---|
-非ポリマー , 5種, 8分子 




| #5: 化合物 |  分子量: 39.098 Da / 分子数: 2 / 由来タイプ: 合成 / 式: K / タイプ: SUBJECT OF INVESTIGATION 分子量: 39.098 Da / 分子数: 2 / 由来タイプ: 合成 / 式: K / タイプ: SUBJECT OF INVESTIGATION#6: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#7: 化合物 | ChemComp-ADP / |  分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM#8: 化合物 | ChemComp-ATP / |  分子量: 507.181 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM#9: 化合物 |  分子量: 54.938 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mn / タイプ: SUBJECT OF INVESTIGATION 分子量: 54.938 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mn / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 283.15 K 詳細: 3UL OF SAMPLE 10C 100% HUMIDITY 30S WAIT TIME 3S BLOT TIME -2 BLOT FORCE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 1800 nm / 最小 デフォーカス(公称値): 800 nm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 69 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 実像数: 4160 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.20.1_4487: / 分類: 精密化 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 3730385 / 詳細: Gaussian blob particle picking in cryoSPARC. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 522000 / アルゴリズム: FOURIER SPACE 詳細: Three different maps were used for final composite map: Hsp90:Cdc37: 288k particles, 3.2A PP5 TPR: 215k particles, 3.3A PP5 catalytic domain and CRaf kinase C-lobe: 22k particles, 4.0A ...詳細: Three different maps were used for final composite map: Hsp90:Cdc37: 288k particles, 3.2A PP5 TPR: 215k particles, 3.3A PP5 catalytic domain and CRaf kinase C-lobe: 22k particles, 4.0A Composite half maps were used to get the final resolution in Relion PostProcessing. All original maps provided in the "Related entries" tab. クラス平均像の数: 3 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT 詳細: This model was built using rigid body docking in Chimera and ChimeraX for all main chains, and RosettaCM to add remaining fragments not included in previous models. Refinement was done using ...詳細: This model was built using rigid body docking in Chimera and ChimeraX for all main chains, and RosettaCM to add remaining fragments not included in previous models. Refinement was done using iterative Phenix and RosettaRelax, and finalized with ISOLDE. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 最高解像度: 3.3 Å |