 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7s06 | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM structure of human GlcNAc-1-phosphotransferase A2B2 subcomplex | ||||||
 要素 要素 | N-acetylglucosamine-1-phosphotransferase subunits alpha/beta | ||||||
 キーワード キーワード | TRANSFERASE / GlcNAc-1-phosphotransferase / lysosomal hydrolases / mannose 6-phosphate trafficking pathway | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報UDP-N-acetylglucosamine-lysosomal-enzyme N-acetylglucosaminephosphotransferase / N-glycan processing to lysosome / secretion of lysosomal enzymes / UDP-N-acetylglucosamine-lysosomal-enzyme N-acetylglucosaminephosphotransferase activity / carbohydrate phosphorylation / lysosome organization / Golgi membrane / calcium ion binding / Golgi apparatus 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.3 Å | ||||||
 データ登録者 データ登録者 | Li, H. / Li, H. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2022 タイトル: Structure of the human GlcNAc-1-phosphotransferase αβ subunits reveals regulatory mechanism for lysosomal enzyme glycan phosphorylation. 著者: Hua Li / Wang-Sik Lee / Xiang Feng / Lin Bai / Benjamin C Jennings / Lin Liu / Balraj Doray / William M Canfield / Stuart Kornfeld / Huilin Li /  要旨: Vertebrates use the mannose 6-phosphate (M6P)-recognition system to deliver lysosomal hydrolases to lysosomes. Key to this pathway is N-acetylglucosamine (GlcNAc)-1-phosphotransferase (PTase) that ...Vertebrates use the mannose 6-phosphate (M6P)-recognition system to deliver lysosomal hydrolases to lysosomes. Key to this pathway is N-acetylglucosamine (GlcNAc)-1-phosphotransferase (PTase) that selectively adds GlcNAc-phosphate (P) to mannose residues of hydrolases. Human PTase is an αβγ heterohexamer with a catalytic core and several peripheral domains that recognize and bind substrates. Here we report a cryo-EM structure of the catalytic core of human PTase and the identification of a hockey stick-like motif that controls activation of the enzyme. Movement of this motif out of the catalytic pocket is associated with a rearrangement of part of the peripheral domains that unblocks hydrolase glycan access to the catalytic site, thereby activating PTase. We propose that PTase fluctuates between inactive and active states in solution, and selective substrate binding of a lysosomal hydrolase through its protein-binding determinant to PTase locks the enzyme in the active state to permit glycan phosphorylation. This mechanism would help ensure that only N-linked glycans of lysosomal enzymes are phosphorylated. #1: ジャーナル: Nat Struct Mol Biol / 年: 2022 タイトル: Bound nucleotide can control the dynamic architecture of monomeric actin. 著者: Rustam Ali / Jacob A Zahm / Michael K Rosen / 要旨: Polymerization of actin into cytoskeletal filaments is coupled to its bound adenine nucleotides. The mechanism by which nucleotide modulates actin functions has not been evident from analyses of ATP- ...Polymerization of actin into cytoskeletal filaments is coupled to its bound adenine nucleotides. The mechanism by which nucleotide modulates actin functions has not been evident from analyses of ATP- and ADP-bound crystal structures of the actin monomer. We report that NMR chemical shift differences between the two forms are globally distributed. Furthermore, microsecond-millisecond motions are spread throughout the molecule in the ATP form, but largely confined to subdomains 1 and 2, and the nucleotide binding site in the ADP form. Through these motions, the ATP- and ADP-bound forms sample different high-energy conformations. A deafness-causing, fast-nucleating actin mutant populates the high-energy conformer of ATP-actin more than the wild-type protein, suggesting that this conformer may be on the pathway to nucleation. Together, the data suggest a model in which differential sampling of a nucleation-compatible form of the actin monomer may contribute to control of actin filament dynamics by nucleotide. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7s06.cif.gz | 207.8 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7s06.ent.gz | 156.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7s06.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7s06_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7s06_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 7s06_validation.xml.gz | 41.4 KB | 表示 | |
| CIF形式データ | 7s06_validation.cif.gz | 60.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/s0/7s06ftp://data.pdbj.org/pub/pdb/validation_reports/s0/7s06 | HTTPS FTP |
-関連構造データ



-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 134787.500 Da / 分子数: 2 / 断片: UNP residues 44-1209 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: GNPTAB, GNPTA, KIAA1208発現宿主:   Cricetulus griseus (モンゴルキヌゲネズミ) Cricetulus griseus (モンゴルキヌゲネズミ)参照: UniProt: Q3T906, UDP-N-acetylglucosamine-lysosomal-enzyme N-acetylglucosaminephosphotransferase #2: 多糖 | 2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose #3: 糖 | ChemComp-NAG / 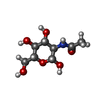  タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C8H15NO6 / タイプ: SUBJECT OF INVESTIGATION タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C8H15NO6 / タイプ: SUBJECT OF INVESTIGATION研究の焦点であるリガンドがあるか | Y | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: GlcNAc-1-phosphotransferase / タイプ: COMPLEX / Entity ID: #1 / 由来: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) | ||||||||||||||||||||
| 由来(組換発現) | 生物種: Cricetulus griseus (モンゴルキヌゲネズミ) | ||||||||||||||||||||
| 緩衝液 | pH: 7.8 | ||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||
| 試料 | 濃度: 0.15 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/1 | ||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 95 % / 凍結前の試料温度: 299 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 105000 X / Cs: 2.7 mm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最高温度: 193 K / 最低温度: 193 K / Residual tilt: 0.05 mradians |
| 撮影 | 平均露光時間: 1.5 sec. / 電子線照射量: 66 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 13320 |
| 画像スキャン | 横: 5760 / 縦: 4092 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 5186047 | ||||||||||||||||
| 3次元再構成 | 解像度: 3.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 66173 / 対称性のタイプ: POINT | ||||||||||||||||
| 原子モデル構築 | プロトコル: AB INITIO MODEL |
