 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7nvm | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
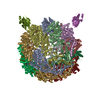
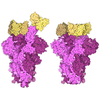
| タイトル | Human TRiC complex in closed state with nanobody Nb18, actin and PhLP2A bound | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | CHAPERONE / TRiC / CCT / ATP hydrolysis / type II chaperonin / protein folding / actin / Structural Genomics / Structural Genomics Consortium / SGC | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報basal body patch / negative regulation of protein folding / perinucleolar compartment / tight junction assembly / zona pellucida receptor complex / positive regulation of establishment of protein localization to telomere / positive regulation of protein localization to Cajal body / profilin binding / scaRNA localization to Cajal body / positive regulation of telomerase RNA localization to Cajal body ...basal body patch / negative regulation of protein folding / perinucleolar compartment / tight junction assembly / zona pellucida receptor complex / positive regulation of establishment of protein localization to telomere / positive regulation of protein localization to Cajal body / profilin binding / scaRNA localization to Cajal body / positive regulation of telomerase RNA localization to Cajal body / chaperonin-containing T-complex / protein localization to bicellular tight junction / BBSome-mediated cargo-targeting to cilium / tubulin complex assembly / regulation of transepithelial transport / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / Gap junction degradation / Formation of tubulin folding intermediates by CCT/TriC / vascular endothelial growth factor receptor 2 binding / binding of sperm to zona pellucida / Cell-extracellular matrix interactions / dense body / Folding of actin by CCT/TriC / regulation of stress fiber assembly / Regulation of CDH1 Function / Prefoldin mediated transfer of substrate to CCT/TriC / Adherens junctions interactions / Sensory processing of sound by outer hair cells of the cochlea / Interaction between L1 and Ankyrins / RHOBTB1 GTPase cycle / regulation of peptidyl-tyrosine phosphorylation / regulation of focal adhesion assembly / Sensory processing of sound by inner hair cells of the cochlea / sarcomere organization / apical junction complex / positive regulation of wound healing / WD40-repeat domain binding / maintenance of blood-brain barrier / myofibril / NuA4 histone acetyltransferase complex / pericentriolar material / filamentous actin / Association of TriC/CCT with target proteins during biosynthesis / Recycling pathway of L1 / sperm head-tail coupling apparatus / EPH-ephrin mediated repulsion of cells / RHO GTPases Activate WASPs and WAVEs / regulation of synaptic vesicle endocytosis / negative regulation of ubiquitin-dependent protein catabolic process / chaperone-mediated protein complex assembly / RHOBTB2 GTPase cycle / RHO GTPases activate IQGAPs / 加水分解酵素; 酸無水物に作用; リン含有酸無水物に作用 / beta-tubulin binding / phagocytic vesicle / heterochromatin / positive regulation of telomere maintenance via telomerase / positive regulation of endothelial cell proliferation / EPHB-mediated forward signaling / axonogenesis / protein folding chaperone / Gene and protein expression by JAK-STAT signaling after Interleukin-12 stimulation / calyx of Held / acrosomal vesicle / FCGR3A-mediated phagocytosis / Translocation of SLC2A4 (GLUT4) to the plasma membrane / actin filament / mRNA 3'-UTR binding / cell motility / ATP-dependent protein folding chaperone / RHO GTPases Activate Formins / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / Regulation of actin dynamics for phagocytic cup formation / VEGFA-VEGFR2 Pathway / structural constituent of cytoskeleton / platelet aggregation / cellular response to type II interferon / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / mRNA 5'-UTR binding / Schaffer collateral - CA1 synapse / response to virus / positive regulation of angiogenesis / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / cell-cell junction / azurophil granule lumen / Signaling by BRAF and RAF1 fusions / melanosome / : / Cooperation of PDCL (PhLP1) and TRiC/CCT in G-protein beta folding / G-protein beta-subunit binding / Clathrin-mediated endocytosis / actin cytoskeleton / protein folding 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.1 Å | |||||||||
 データ登録者 データ登録者 | Kelly, J.J. / Chi, G. / Bulawa, C. / Paavilainen, V.O. / Bountra, C. / Huiskonen, J.T. / Yue, W. / Structural Genomics Consortium (SGC) | |||||||||
| 資金援助 |  英国, 英国,  フィンランド, 2件 フィンランド, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2022 タイトル: Snapshots of actin and tubulin folding inside the TRiC chaperonin. 著者: John J Kelly / Dale Tranter / Els Pardon / Gamma Chi / Holger Kramer / Lotta Happonen / Kelly M Knee / Jay M Janz / Jan Steyaert / Christine Bulawa / Ville O Paavilainen / Juha T Huiskonen / Wyatt W Yue /    要旨: The integrity of a cell's proteome depends on correct folding of polypeptides by chaperonins. The chaperonin TCP-1 ring complex (TRiC) acts as obligate folder for >10% of cytosolic proteins, ...The integrity of a cell's proteome depends on correct folding of polypeptides by chaperonins. The chaperonin TCP-1 ring complex (TRiC) acts as obligate folder for >10% of cytosolic proteins, including he cytoskeletal proteins actin and tubulin. Although its architecture and how it recognizes folding substrates are emerging from structural studies, the subsequent fate of substrates inside the TRiC chamber is not defined. We trapped endogenous human TRiC with substrates (actin, tubulin) and cochaperone (PhLP2A) at different folding stages, for structure determination by cryo-EM. The already-folded regions of client proteins are anchored at the chamber wall, positioning unstructured regions toward the central space to achieve their native fold. Substrates engage with different sections of the chamber during the folding cycle, coupled to TRiC open-and-close transitions. Further, the cochaperone PhLP2A modulates folding, acting as a molecular strut between substrate and TRiC chamber. Our structural snapshots piece together an emerging model of client protein folding within TRiC. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7nvm.cif.gz | 1.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7nvm.ent.gz | 1.2 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7nvm.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/nv/7nvmftp://data.pdbj.org/pub/pdb/validation_reports/nv/7nvm | HTTPS FTP |
|---|
-関連構造データ








-リンク
 PDBj
PDBj



























- 集合体
集合体
| 登録構造単位 | 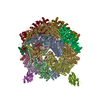
|
|---|---|
| 1 |
|
-要素
-T-complex protein 1 subunit ... , 8種, 16分子 AaBbDdEeGgHhQqZz
| #1: タンパク質 | 分子量: 60418.477 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P17987#2: タンパク質 | 分子量: 57567.141 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P78371#3: タンパク質 | 分子量: 57996.113 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P50991#4: タンパク質 | 分子量: 59749.957 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CCT5, CCTE, KIAA0098 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P48643#5: タンパク質 | 分子量: 60613.855 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CCT3, CCTG, TRIC5 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P49368#6: タンパク質 | 分子量: 59443.535 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q99832#8: タンパク質 | 分子量: 59691.422 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P50990#9: タンパク質 | 分子量: 58106.086 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P40227 |
|---|
-タンパク質 , 2種, 2分子 KP
| #10: タンパク質 | 分子量: 41838.766 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P63261 |
|---|---|
| #11: タンパク質 | 分子量: 27650.383 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q9H2J4 |
-抗体 , 1種, 2分子 Nn
| #7: 抗体 | 分子量: 14412.816 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現)  |
|---|
-非ポリマー , 4種, 67分子 

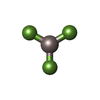

| #12: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 16 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 16 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM#13: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 16 / 由来タイプ: 天然 / 式: Mg 分子量: 24.305 Da / 分子数: 16 / 由来タイプ: 天然 / 式: Mg#14: 化合物 | ChemComp-AF3 /  分子量: 83.977 Da / 分子数: 16 / 由来タイプ: 合成 / 式: AlF3 分子量: 83.977 Da / 分子数: 16 / 由来タイプ: 合成 / 式: AlF3#15: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 19 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 43 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.1 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 63082 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 71.38 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
