 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6sb0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
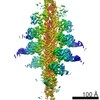
| タイトル | cryo-EM structure of mTORC1 bound to PRAS40-fused active RagA/C GTPases | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | SIGNALING PROTEIN / small GTPases / mTORC1 activator / roadblock domain / GTPase domain | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Gtr1-Gtr2 GTPase complex / FNIP-folliculin RagC/D GAP / RNA polymerase III type 2 promoter sequence-specific DNA binding / RNA polymerase III type 1 promoter sequence-specific DNA binding / positive regulation of cytoplasmic translational initiation / regulation of locomotor rhythm / T-helper 1 cell lineage commitment / positive regulation of pentose-phosphate shunt / positive regulation of wound healing, spreading of epidermal cells / TORC2 complex ...Gtr1-Gtr2 GTPase complex / FNIP-folliculin RagC/D GAP / RNA polymerase III type 2 promoter sequence-specific DNA binding / RNA polymerase III type 1 promoter sequence-specific DNA binding / positive regulation of cytoplasmic translational initiation / regulation of locomotor rhythm / T-helper 1 cell lineage commitment / positive regulation of pentose-phosphate shunt / positive regulation of wound healing, spreading of epidermal cells / TORC2 complex / cellular response to leucine starvation / TFIIIC-class transcription factor complex binding / regulation of TORC1 signaling / regulation of membrane permeability / negative regulation of lysosome organization / heart valve morphogenesis / nucleus localization / protein localization to lysosome / RNA polymerase III type 3 promoter sequence-specific DNA binding / TORC1 complex / positive regulation of transcription of nucleolar large rRNA by RNA polymerase I / regulation of TOR signaling / calcineurin-NFAT signaling cascade / voluntary musculoskeletal movement / TORC1 signaling / regulation of osteoclast differentiation / positive regulation of odontoblast differentiation / positive regulation of keratinocyte migration / cellular response to L-leucine / Amino acids regulate mTORC1 / MTOR signalling / cellular response to nutrient / regulation of autophagosome assembly / cellular response to methionine / Energy dependent regulation of mTOR by LKB1-AMPK / energy reserve metabolic process / negative regulation of cell size / protein localization to membrane / ruffle organization / positive regulation of osteoclast differentiation / cellular response to osmotic stress / protein serine/threonine kinase inhibitor activity / negative regulation of TOR signaling / enzyme-substrate adaptor activity / negative regulation of protein localization to nucleus / anoikis / cardiac muscle cell development / AKT phosphorylates targets in the cytosol / positive regulation of transcription by RNA polymerase III / negative regulation of calcineurin-NFAT signaling cascade / regulation of myelination / positive regulation of actin filament polymerization / small GTPase-mediated signal transduction / regulation of cell size / negative regulation of macroautophagy / positive regulation of oligodendrocyte differentiation / lysosome organization / Macroautophagy / positive regulation of myotube differentiation / protein kinase activator activity / behavioral response to pain / protein kinase inhibitor activity / oligodendrocyte differentiation / Constitutive Signaling by AKT1 E17K in Cancer / mTORC1-mediated signalling / germ cell development / : / CD28 dependent PI3K/Akt signaling / neurotrophin TRK receptor signaling pathway / social behavior / HSF1-dependent transactivation / neuronal action potential / positive regulation of TOR signaling / response to amino acid / TOR signaling / endomembrane system / 'de novo' pyrimidine nucleobase biosynthetic process / regulation of macroautophagy / positive regulation of translational initiation / positive regulation of G1/S transition of mitotic cell cycle / cellular response to nutrient levels / regulation of neuron apoptotic process / positive regulation of lamellipodium assembly / phagocytic vesicle / heart morphogenesis / positive regulation of lipid biosynthetic process / positive regulation of epithelial to mesenchymal transition / phosphorylation / cardiac muscle contraction / regulation of cellular response to heat / protein-membrane adaptor activity / positive regulation of stress fiber assembly / tumor necrosis factor-mediated signaling pathway / negative regulation of TORC1 signaling / cytoskeleton organization / 14-3-3 protein binding / positive regulation of endothelial cell proliferation / T cell costimulation / positive regulation of TORC1 signaling / cellular response to amino acid starvation 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 5.5 Å | ||||||||||||
 データ登録者 データ登録者 | Anandapadamanaban, M. / Berndt, A. / Masson, G.R. / Perisic, O. / Williams, R.L. | ||||||||||||
| 資金援助 |  英国, 3件 英国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Science / 年: 2019 タイトル: Architecture of human Rag GTPase heterodimers and their complex with mTORC1. 著者: Madhanagopal Anandapadamanaban / Glenn R Masson / Olga Perisic / Alex Berndt / Jonathan Kaufman / Chris M Johnson / Balaji Santhanam / Kacper B Rogala / David M Sabatini / Roger L Williams /  要旨: The Rag guanosine triphosphatases (GTPases) recruit the master kinase mTORC1 to lysosomes to regulate cell growth and proliferation in response to amino acid availability. The nucleotide state of Rag ...The Rag guanosine triphosphatases (GTPases) recruit the master kinase mTORC1 to lysosomes to regulate cell growth and proliferation in response to amino acid availability. The nucleotide state of Rag heterodimers is critical for their association with mTORC1. Our cryo-electron microscopy structure of RagA/RagC in complex with mTORC1 shows the details of RagA/RagC binding to the RAPTOR subunit of mTORC1 and explains why only the RagA/RagC nucleotide state binds mTORC1. Previous kinetic studies suggested that GTP binding to one Rag locks the heterodimer to prevent GTP binding to the other. Our crystal structures and dynamics of RagA/RagC show the mechanism for this locking and explain how oncogenic hotspot mutations disrupt this process. In contrast to allosteric activation by RHEB, Rag heterodimer binding does not change mTORC1 conformation and activates mTORC1 by targeting it to lysosomes. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6sb0.cif.gz | 1.2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6sb0.ent.gz | 867.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6sb0.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6sb0_validation.pdf.gz | 1.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6sb0_full_validation.pdf.gz | 1.6 MB | 表示 | |
| XML形式データ | 6sb0_validation.xml.gz | 167.5 KB | 表示 | |
| CIF形式データ | 6sb0_validation.cif.gz | 286.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/sb/6sb0ftp://data.pdbj.org/pub/pdb/validation_reports/sb/6sb0 | HTTPS FTP |
-関連構造データ







-リンク
 PDBj
PDBj










- 集合体
集合体
| 登録構造単位 | 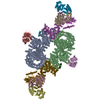
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 非結晶学的対称性 (NCS) | NCSドメイン:
NCSドメイン領域:
NCSアンサンブル:
NCS oper:
|
-要素
-タンパク質 , 4種, 8分子 ABEHYNTO
| #1: タンパク質 | 分子量: 287012.031 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MTOR, FRAP, FRAP1, FRAP2, RAFT1, RAPT1 / 発現宿主: Homo sapiens (ヒト)参照: UniProt: P42345, non-specific serine/threonine protein kinase #2: タンパク質 | 分子量: 35910.090 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MLST8, GBL, LST8 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q9BVC4#5: タンパク質 | 分子量: 149200.016 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RPTOR, KIAA1303, RAPTOR / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q8N122#6: タンパク質 | 分子量: 27412.293 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: AKT1S1, PRAS40 / 発現宿主:  |
|---|
-Ras-related GTP-binding protein ... , 2種, 4分子 CIDJ
| #3: タンパク質 | 分子量: 36600.195 Da / 分子数: 2 / Mutation: Q66L / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGA / 発現宿主: #4: タンパク質 | 分子量: 44284.832 Da / 分子数: 2 / Mutation: T90N / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGC / 発現宿主: |
|---|
-非ポリマー , 2種, 4分子 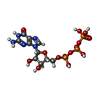
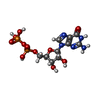
| #7: 化合物 |  分子量: 523.180 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: GTP, エネルギー貯蔵分子*YM 分子量: 523.180 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: GTP, エネルギー貯蔵分子*YM#8: 化合物 |  タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: GDP, エネルギー貯蔵分子*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 1.09 MDa / 実験値: YES | ||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||
| 緩衝液 | pH: 7 / 詳細: 50mM HEPES pH 7.0, 100mM NaCl, 2mM MgCl2, 1mM TCEP | ||||||||||||||||||||||||
| 試料 | 濃度: 0.05 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: mTORC1 (mTOR complex 1) is a dimer consists of three proteins: mTOR, mLST8 and RAPTOR. The interacting partner, PRAS40-fused-RagA/C (referred as RagA/C here) forms complex with mTORC1 for its ...詳細: mTORC1 (mTOR complex 1) is a dimer consists of three proteins: mTOR, mLST8 and RAPTOR. The interacting partner, PRAS40-fused-RagA/C (referred as RagA/C here) forms complex with mTORC1 for its activation. We solved the cryo-EM structure of mTORC1 bound to RagA/C. | ||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE / 湿度: 95 % |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / Cs: 2.7 mm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.8 sec. / 電子線照射量: 40 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
| 画像スキャン | 横: 3838 / 縦: 3710 / 動画フレーム数/画像: 22 / 利用したフレーム数/画像: 1-22 |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0238 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画像処理 | 詳細: The selected images were processed using MotionCor2 within the RELION-3.0.6 package. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 580707 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 5.5 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 90809 / アルゴリズム: FOURIER SPACE 詳細: For the final reconstruction of mTORC1-RagA/C structure we used a strategy taking advantage of the relion particle symmetry expand program, and duplicated the C2-refined particles and applied ...詳細: For the final reconstruction of mTORC1-RagA/C structure we used a strategy taking advantage of the relion particle symmetry expand program, and duplicated the C2-refined particles and applied the appropriate rotation and translation to generate a set of monomers. We performed mTORC1-RagA/C 'pseudo-monomer' focussed classification with signal subtraction and obtained a reconstruction of 5.5 A resolution map. This cryo-EM density corresponded to the mTORC1-RagA/C pseudomonomer, where the previously published structure for apo-mTORC1 (PDB ID 6BCX) and our high-resolution crystal structure of RagA/C (6S6A) were fitted with great confidence from our experimental analysis including Pulldown assays, mutational at per-residue level in the binding interface and HDX-Mass Spectrometry. クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 283 / プロトコル: RIGID BODY FIT / 空間: REAL 詳細: Cryo-EM model of mTORC1-RagA/C was refined using REFMAC5 program in CCPEM package, with a composite map of the 3D reconstruction of mTORC1-RagA/C pseudo-monomer (as mentioned in ...詳細: Cryo-EM model of mTORC1-RagA/C was refined using REFMAC5 program in CCPEM package, with a composite map of the 3D reconstruction of mTORC1-RagA/C pseudo-monomer (as mentioned in Reconstruction section) of one protomer together with the generated map for the other second protomer of mTORC1-RagA/C. This second protomer of mTORC1-RagA/C map was generated by simply aligning the first 3D reconstructed pseudomonomer map onto the mTORC1 dimer consensus C2 map and then obtained the rotation-translation matrix with CHIMERA and then used Maputils program in CCP4i. From the resulting mTORC1-RagA/C dimer map, the model of mTORC1-RagA/C was built by using previously published structure of apo-mTORC1 (PDB ID 6BCX) and our crystal structure of RagA/C was fitted (PDB ID 6S6A, unreleased). The entire mTORC1-RagA/C final model was refined using REFMAC5 program using the restraints from the crystal structure of RagA/C and previously published mTORC1 structure. Side chains were removed before refinement, since these were not evident in the cryo-EM densities. Separate model refinements were performed against single half-maps, and the resulting models were compared with the other half-maps to confirm the absence of overfitting. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 5.5→321.75 Å / Cor.coef. Fo:Fc: 0.895 / SU B: 104.924 / SU ML: 1.049 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 溶媒モデル: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 288.4 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 66022 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
