Stealth protein CR2, conserved region 2 / Stealth protein CR4, conserved region 4 / Stealth protein CR3, conserved region 3 / Stealth protein CR1, conserved region 1 / Stealth protein CR2, conserved region 2 / Stealth protein CR1, conserved region 1 / Stealth protein CR3, conserved region 3 / Stealth protein CR4, conserved region 4 / Notch-like domain superfamily / LNR (Lin-12/Notch) repeat profile. ...Stealth protein CR2, conserved region 2 / Stealth protein CR4, conserved region 4 / Stealth protein CR3, conserved region 3 / Stealth protein CR1, conserved region 1 / Stealth protein CR2, conserved region 2 / Stealth protein CR1, conserved region 1 / Stealth protein CR3, conserved region 3 / Stealth protein CR4, conserved region 4 / Notch-like domain superfamily / LNR (Lin-12/Notch) repeat profile. / LNR domain / Notch domain / Domain found in Notch and Lin-12 類似検索 - ドメイン・相同性
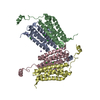
ジャーナル: J Biol Chem / 年: 2022 タイトル: Structural insights into how GlcNAc-1-phosphotransferase directs lysosomal protein transport. 著者: Shuo Du / Guopeng Wang / Zhiying Zhang / Chengying Ma / Ning Gao / Junyu Xiao / 要旨: GlcNAc-1-phosphotransferase catalyzes the initial step in the formation of the mannose-6-phosphate tag that labels ∼60 lysosomal proteins for transport. Mutations in GlcNAc-1-phosphotransferase are ...GlcNAc-1-phosphotransferase catalyzes the initial step in the formation of the mannose-6-phosphate tag that labels ∼60 lysosomal proteins for transport. Mutations in GlcNAc-1-phosphotransferase are known to cause lysosomal storage disorders such as mucolipidoses. However, the molecular mechanism of GlcNAc-1-phosphotransferase activity remains unclear. Mammalian GlcNAc-1-phosphotransferases are α2β2γ2 hexamers in which the core catalytic α- and β-subunits are derived from the GNPTAB (N-acetylglucosamine-1-phosphate transferase subunits alpha and beta) gene. Here, we present the cryo-electron microscopy structure of the Drosophila melanogaster GNPTAB homolog, DmGNPTAB. We identified four conserved regions located far apart in the sequence that fold into the catalytic domain, which exhibits structural similarity to that of the UDP-glucose glycoprotein glucosyltransferase. Comparison with UDP-glucose glycoprotein glucosyltransferase also revealed a putative donor substrate-binding site, and the functional requirements of critical residues in human GNPTAB were validated using GNPTAB-knockout cells. Finally, we show that DmGNPTAB forms a homodimer that is evolutionarily conserved and that perturbing the dimer interface undermines the maturation and activity of human GNPTAB. These results provide important insights into GlcNAc-1-phosphotransferase function and related diseases.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用
 構造の表示
構造の表示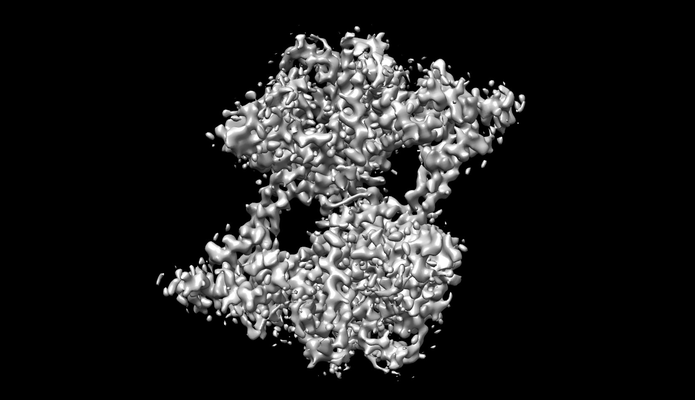
 ダウンロードとリンク
ダウンロードとリンク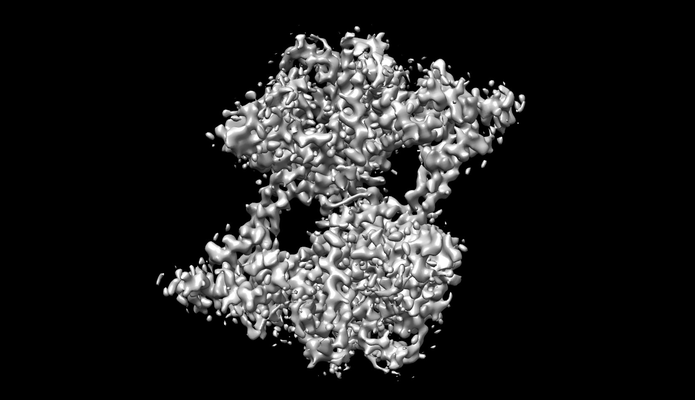 emd_30910.png
emd_30910.png http://ftp.pdbj.org/pub/emdb/structures/EMD-30910
http://ftp.pdbj.org/pub/emdb/structures/EMD-30910




 Z
Z Y
Y X
X







 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
