Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
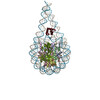
| Title | Trinucleosome of the 4x197 nucleosome array containing H1 | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | Chromatin / Nucleosome / Linker Histone / DNA BINDING PROTEIN | |||||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of DNA recombination / Apoptosis induced DNA fragmentation / chromosome condensation / nucleosomal DNA binding / Formation of Senescence-Associated Heterochromatin Foci (SAHF) / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / heterochromatin ...negative regulation of DNA recombination / Apoptosis induced DNA fragmentation / chromosome condensation / nucleosomal DNA binding / Formation of Senescence-Associated Heterochromatin Foci (SAHF) / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / heterochromatin / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / Interleukin-7 signaling / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / innate immune response in mucosa / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / euchromatin / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / HDMs demethylate histones / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / chromatin DNA binding / G2/M DNA damage checkpoint / Negative Regulation of CDH1 Gene Transcription / NoRC negatively regulates rRNA expression / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / Meiotic recombination / Pre-NOTCH Transcription and Translation / Activation of anterior HOX genes in hindbrain development during early embryogenesis / histone deacetylase binding / Transcriptional regulation of granulopoiesis / RMTs methylate histone arginines / Metalloprotease DUBs / HCMV Early Events / structural constituent of chromatin / UCH proteinases / nucleosome / antimicrobial humoral immune response mediated by antimicrobial peptide / heterochromatin formation / nucleosome assembly / HATs acetylate histones / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / Factors involved in megakaryocyte development and platelet production / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / chromatin organization / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / double-stranded DNA binding / Oxidative Stress Induced Senescence / Estrogen-dependent gene expression / killing of cells of another organism / gene expression / defense response to Gram-negative bacterium / chromosome, telomeric region / Ub-specific processing proteases / defense response to Gram-positive bacterium / Amyloid fiber formation / protein heterodimerization activity / negative regulation of cell population proliferation / chromatin binding / negative regulation of transcription by RNA polymerase II / protein-containing complex / : / DNA binding / RNA binding / extracellular exosome / extracellular region / nucleoplasm / membrane / nucleus / cytosol Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) / synthetic construct (others) Homo sapiens (human) / synthetic construct (others) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 9.7 Å | |||||||||
 Authors Authors | Dombrowski M / Cramer P | |||||||||
| Funding support | European Union, 1 items
| |||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2022 Title: Histone H1 binding to nucleosome arrays depends on linker DNA length and trajectory. Authors: Marco Dombrowski / Maik Engeholm / Christian Dienemann / Svetlana Dodonova / Patrick Cramer /  Abstract: Throughout the genome, nucleosomes often form regular arrays that differ in nucleosome repeat length (NRL), occupancy of linker histone H1 and transcriptional activity. Here, we report cryo-EM ...Throughout the genome, nucleosomes often form regular arrays that differ in nucleosome repeat length (NRL), occupancy of linker histone H1 and transcriptional activity. Here, we report cryo-EM structures of human H1-containing tetranucleosome arrays with four physiologically relevant NRLs. The structures show a zig-zag arrangement of nucleosomes, with nucleosomes 1 and 3 forming a stack. H1 binding to stacked nucleosomes depends on the NRL, whereas H1 always binds to the non-stacked nucleosomes 2 and 4. Short NRLs lead to altered trajectories of linker DNA, and these altered trajectories sterically impair H1 binding to the stacked nucleosomes in our structures. As the NRL increases, linker DNA trajectories relax, enabling H1 contacts and binding. Our results provide an explanation for why arrays with short NRLs are depleted of H1 and suited for transcription, whereas arrays with long NRLs show full H1 occupancy and can form transcriptionally silent heterochromatin regions. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_13370.map.gz | 16.2 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-13370-v30.xmlemd-13370.xml | 21.2 KB 21.2 KB | Display Display | EMDB header |
| FSC (resolution estimation) | emd_13370_fsc.xml | 7.2 KB | Display | FSC data file |
| Images |  emd_13370.png emd_13370.png | 22 KB | ||
| Filedesc metadata | emd-13370.cif.gz | 6.4 KB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-13370ftp://ftp.pdbj.org/pub/emdb/structures/EMD-13370 http://ftp.pdbj.org/pub/emdb/structures/EMD-13370ftp://ftp.pdbj.org/pub/emdb/structures/EMD-13370 | HTTPS FTP |
-Related structure data
| Related structure data |  7pfaMC  7petC  7peuC  7pevC  7pewC  7pexC  7peyC  7pezC  7pf0C  7pf2C  7pf3C  7pf4C  7pf5C  7pf6C  7pfcC  7pfdC  7pfeC  7pffC  7pftC  7pfuC  7pfvC  7pfwC  7pfxC M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |




















-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |















-Map
| File | Download / File: emd_13370.map.gz / Format: CCP4 / Size: 30.5 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 3.15 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
- Sample components
Sample components
-Entire : Trinucleosome from 4x197 array
| Entire | Name: Trinucleosome from 4x197 array |
|---|---|
| Components |
|
-Supramolecule #1: Trinucleosome from 4x197 array
| Supramolecule | Name: Trinucleosome from 4x197 array / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all |
|---|
-Macromolecule #1: Histone H3.2
| Macromolecule | Name: Histone H3.2 / type: protein_or_peptide / ID: 1 / Number of copies: 6 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 15.389036 KDa |
| Recombinant expression | Organism:  |
| Sequence | String: MARTKQTARK STGGKAPRKQ LATKAARKSA PATGGVKKPH RYRPGTVALR EIRRYQKSTE LLIRKLPFQR LVREIAQDFK TDLRFQSSA VMALQEASEA YLVGLFEDTN LAAIHAKRVT IMPKDIQLAR RIRGERA UniProtKB: Histone H3.2 |
-Macromolecule #2: Histone H4
| Macromolecule | Name: Histone H4 / type: protein_or_peptide / ID: 2 / Number of copies: 6 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 11.394426 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MSGRGKGGKG LGKGGAKRHR KVLRDNIQGI TKPAIRRLAR RGGVKRISGL IYEETRGVLK VFLENVIRDA VTYTEHAKRK TVTAMDVVY ALKRQGRTLY GFGG UniProtKB: Histone H4 |
-Macromolecule #3: Histone H2A type 1-B/E
| Macromolecule | Name: Histone H2A type 1-B/E / type: protein_or_peptide / ID: 3 / Number of copies: 6 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 16.344873 KDa |
| Recombinant expression | Organism: |
| Sequence | String: HHHHHHENLY FQSNAPWMSG RGKQGGKARA KAKTRSSRAG LQFPVGRVHR LLRKGNYSER VGAGAPVYLA AVLEYLTAEI LELAGNAAR DNKKTRIIPR HLQLAIRNDE ELNKLLGRVT IAQGGVLPNI QAVLLPKKTE SHHKAKGK UniProtKB: Histone H2A type 1-B/E |
-Macromolecule #4: Histone H2B type 1-K
| Macromolecule | Name: Histone H2B type 1-K / type: protein_or_peptide / ID: 4 / Number of copies: 6 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 13.921213 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MPEPAKSAPA PKKGSKKAVT KAQKKDGKKR KRSRKESYSV YVYKVLKQVH PDTGISSKAM GIMNSFVNDI FERIAGEASR LAHYNKRST ITSREIQTAV RLLLPGELAK HAVSEGTKAV TKYTSAK UniProtKB: Histone H2B type 1-K |
-Macromolecule #5: Histone H1.4
| Macromolecule | Name: Histone H1.4 / type: protein_or_peptide / ID: 5 / Number of copies: 2 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 21.800326 KDa |
| Recombinant expression | Organism: |
| Sequence | String: SETAPAAPAA PAPAEKTPVK KKARKSAGAA KRKASGPPVS ELITKAVAAS KERSGVSLAA LKKALAAAGY DVEKNNSRIK LGLKSLVSK GTLVQTKGTG ASGSFKLNKK AASGEAKPKA KKAGAAKAKK PAGAAKKPKK ATGAATPKKS AKKTPKKAKK P AAAAGAKK ...String: SETAPAAPAA PAPAEKTPVK KKARKSAGAA KRKASGPPVS ELITKAVAAS KERSGVSLAA LKKALAAAGY DVEKNNSRIK LGLKSLVSK GTLVQTKGTG ASGSFKLNKK AASGEAKPKA KKAGAAKAKK PAGAAKKPKK ATGAATPKKS AKKTPKKAKK P AAAAGAKK AKSPKKAKAA KPKKAPKSPA KAKAVKPKAA KPKTAKPKAA KPKKAAAKKK UniProtKB: Histone H1.4 |
-Macromolecule #6: DNA (561-MER)
| Macromolecule | Name: DNA (561-MER) / type: dna / ID: 6 / Number of copies: 1 / Classification: DNA |
|---|---|
| Source (natural) | Organism: synthetic construct (others) |
| Molecular weight | Theoretical: 242.284766 KDa |
| Sequence | String: (DA)(DT)(DC)(DG)(DT)(DC)(DT)(DC)(DG)(DC) (DG)(DC)(DA)(DC)(DT)(DG)(DG)(DC)(DC)(DG) (DC)(DC)(DA)(DT)(DA)(DC)(DT)(DG)(DG) (DA)(DG)(DA)(DA)(DT)(DC)(DC)(DC)(DG)(DG) (DT) (DG)(DC)(DC)(DG)(DA)(DG) ...String: (DA)(DT)(DC)(DG)(DT)(DC)(DT)(DC)(DG)(DC) (DG)(DC)(DA)(DC)(DT)(DG)(DG)(DC)(DC)(DG) (DC)(DC)(DA)(DT)(DA)(DC)(DT)(DG)(DG) (DA)(DG)(DA)(DA)(DT)(DC)(DC)(DC)(DG)(DG) (DT) (DG)(DC)(DC)(DG)(DA)(DG)(DG)(DC) (DC)(DG)(DC)(DT)(DC)(DA)(DA)(DT)(DT)(DG) (DG)(DT) (DC)(DG)(DT)(DA)(DG)(DA)(DC) (DA)(DG)(DC)(DT)(DC)(DT)(DA)(DG)(DC)(DA) (DC)(DC)(DG) (DC)(DT)(DT)(DA)(DA)(DA) (DC)(DG)(DC)(DA)(DC)(DG)(DT)(DA)(DC)(DG) (DC)(DG)(DC)(DT) (DG)(DT)(DC)(DC)(DC) (DC)(DC)(DG)(DC)(DG)(DT)(DT)(DT)(DT)(DA) (DA)(DC)(DC)(DG)(DC) (DC)(DA)(DA)(DG) (DG)(DG)(DG)(DA)(DT)(DT)(DA)(DC)(DT)(DC) (DC)(DC)(DT)(DA)(DG)(DT) (DC)(DT)(DC) (DC)(DA)(DG)(DG)(DC)(DA)(DC)(DG)(DT)(DG) (DT)(DC)(DA)(DG)(DA)(DT)(DA) (DT)(DA) (DT)(DA)(DC)(DA)(DT)(DC)(DC)(DT)(DG)(DT) (DC)(DA)(DT)(DG)(DT)(DA)(DA)(DG) (DT) (DA)(DT)(DT)(DA)(DA)(DG)(DG)(DT)(DA)(DA) (DC)(DC)(DC)(DA)(DG)(DT)(DA)(DC)(DT) (DG)(DT)(DC)(DT)(DC)(DG)(DC)(DG)(DC)(DA) (DC)(DT)(DG)(DG)(DC)(DC)(DG)(DC)(DC)(DA) (DT)(DA)(DC)(DT)(DG)(DG)(DA)(DG)(DA) (DA)(DT)(DC)(DC)(DC)(DG)(DG)(DT)(DG)(DC) (DC) (DG)(DA)(DG)(DG)(DC)(DC)(DG)(DC) (DT)(DC)(DA)(DA)(DT)(DT)(DG)(DG)(DT)(DC) (DG)(DT) (DA)(DG)(DA)(DC)(DA)(DG)(DC) (DT)(DC)(DT)(DA)(DG)(DC)(DA)(DC)(DC)(DG) (DC)(DT)(DT) (DA)(DA)(DA)(DC)(DG)(DC) (DA)(DC)(DG)(DT)(DA)(DC)(DG)(DC)(DG)(DC) (DT)(DG)(DT)(DC) (DC)(DC)(DC)(DC)(DG) (DC)(DG)(DT)(DT)(DT)(DT)(DA)(DA)(DC)(DC) (DG)(DC)(DC)(DA)(DA) (DG)(DG)(DG)(DG) (DA)(DT)(DT)(DA)(DC)(DT)(DC)(DC)(DC)(DT) (DA)(DG)(DT)(DC)(DT)(DC) (DC)(DA)(DG) (DG)(DC)(DA)(DC)(DG)(DT)(DG)(DT)(DC)(DA) (DG)(DA)(DT)(DA)(DT)(DA)(DT) (DA)(DC) (DA)(DT)(DC)(DC)(DT)(DG)(DT)(DC)(DA)(DT) (DG)(DT)(DA)(DA)(DG)(DT)(DA)(DT) (DT) (DA)(DA)(DG)(DG)(DT)(DA)(DA)(DC)(DC)(DC) (DA)(DG)(DT)(DA)(DC)(DT)(DG)(DT)(DC) (DT)(DC)(DG)(DC)(DG)(DC)(DA)(DC)(DT)(DG) (DG)(DC)(DC)(DG)(DC)(DC)(DA)(DT)(DA)(DC) (DT)(DG)(DG)(DA)(DG)(DA)(DA)(DT)(DC) (DC)(DC)(DG)(DG)(DT)(DG)(DC)(DC)(DG)(DA) (DG) (DG)(DC)(DC)(DG)(DC)(DT)(DC)(DA) (DA)(DT)(DT)(DG)(DG)(DT)(DC)(DG)(DT)(DA) (DG)(DA) (DC)(DA)(DG)(DC)(DT)(DC)(DT) (DA)(DG)(DC)(DA)(DC)(DC)(DG)(DC)(DT)(DT) (DA)(DA)(DA) (DC)(DG)(DC)(DA)(DC)(DG) (DT)(DA)(DC)(DG)(DC)(DG)(DC)(DT)(DG)(DT) (DC)(DC)(DC)(DC) (DC)(DG)(DC)(DG)(DT) (DT)(DT)(DT)(DA)(DA)(DC)(DC)(DG)(DC)(DC) (DA)(DA)(DG)(DG)(DG) (DG)(DA)(DT)(DT) (DA)(DC)(DT)(DC)(DC)(DC)(DT)(DA)(DG)(DT) (DC)(DT)(DC)(DC)(DA)(DG) (DG)(DC)(DA) (DC)(DG)(DT)(DG)(DT)(DC)(DA)(DG)(DA)(DT) (DA)(DT)(DA)(DT)(DA)(DC)(DA) (DT)(DC) (DC)(DT)(DG)(DT)(DC)(DA)(DT)(DG)(DT)(DA) (DA)(DG)(DT)(DA)(DT)(DT)(DA)(DA) (DG) (DG)(DT)(DA)(DA)(DC)(DC)(DC)(DA)(DG)(DT) (DA)(DC)(DT)(DG)(DT)(DC)(DT)(DC)(DG) (DC)(DG)(DC)(DA)(DC)(DT)(DG)(DG)(DC)(DC) (DG)(DC)(DC)(DA)(DT)(DA)(DC)(DT)(DG)(DG) (DA)(DG)(DA)(DA)(DT)(DC)(DC)(DC)(DG) (DG)(DT)(DG)(DC)(DC)(DG)(DA)(DG)(DG)(DC) (DC) (DG)(DC)(DT)(DC)(DA)(DA)(DT)(DT) (DG)(DG)(DT)(DC)(DG)(DT)(DA)(DG)(DA)(DC) (DA)(DG) (DC)(DT)(DC)(DT)(DA)(DG)(DC) (DA)(DC)(DC)(DG)(DC)(DT)(DT)(DA)(DA)(DA) (DC)(DG)(DC) (DA)(DC)(DG)(DT)(DA)(DC) (DG)(DC)(DG)(DC)(DT)(DG)(DT)(DC)(DC)(DC) (DC)(DC)(DG)(DC) (DG)(DT)(DT)(DT)(DT) (DA)(DA)(DC)(DC)(DG)(DC)(DC)(DA)(DA)(DG) (DG)(DG)(DG)(DA)(DT) (DT)(DA)(DC)(DT) (DC)(DC)(DC)(DT)(DA)(DG)(DT)(DC)(DT)(DC) (DC)(DA)(DG)(DG)(DC)(DA) (DC)(DG)(DT) (DG)(DT)(DC)(DA)(DG)(DA)(DT)(DA)(DT)(DA) (DT)(DA)(DC)(DA)(DT)(DC)(DC) (DT)(DG) (DT)(DC)(DA)(DT)(DG)(DT)(DA)(DA)(DG)(DT) (DA)(DT)(DT)(DA)(DA)(DG)(DG)(DT) (DA) (DA)(DC)(DC)(DC)(DG)(DA)(DT) |
-Macromolecule #7: DNA (561-MER)
| Macromolecule | Name: DNA (561-MER) / type: dna / ID: 7 / Number of copies: 1 / Classification: DNA |
|---|---|
| Source (natural) | Organism: synthetic construct (others) |
| Molecular weight | Theoretical: 244.561938 KDa |
| Sequence | String: (DA)(DT)(DC)(DG)(DG)(DG)(DT)(DT)(DA)(DC) (DC)(DT)(DT)(DA)(DA)(DT)(DA)(DC)(DT)(DT) (DA)(DC)(DA)(DT)(DG)(DA)(DC)(DA)(DG) (DG)(DA)(DT)(DG)(DT)(DA)(DT)(DA)(DT)(DA) (DT) (DC)(DT)(DG)(DA)(DC)(DA) ...String: (DA)(DT)(DC)(DG)(DG)(DG)(DT)(DT)(DA)(DC) (DC)(DT)(DT)(DA)(DA)(DT)(DA)(DC)(DT)(DT) (DA)(DC)(DA)(DT)(DG)(DA)(DC)(DA)(DG) (DG)(DA)(DT)(DG)(DT)(DA)(DT)(DA)(DT)(DA) (DT) (DC)(DT)(DG)(DA)(DC)(DA)(DC)(DG) (DT)(DG)(DC)(DC)(DT)(DG)(DG)(DA)(DG)(DA) (DC)(DT) (DA)(DG)(DG)(DG)(DA)(DG)(DT) (DA)(DA)(DT)(DC)(DC)(DC)(DC)(DT)(DT)(DG) (DG)(DC)(DG) (DG)(DT)(DT)(DA)(DA)(DA) (DA)(DC)(DG)(DC)(DG)(DG)(DG)(DG)(DG)(DA) (DC)(DA)(DG)(DC) (DG)(DC)(DG)(DT)(DA) (DC)(DG)(DT)(DG)(DC)(DG)(DT)(DT)(DT)(DA) (DA)(DG)(DC)(DG)(DG) (DT)(DG)(DC)(DT) (DA)(DG)(DA)(DG)(DC)(DT)(DG)(DT)(DC)(DT) (DA)(DC)(DG)(DA)(DC)(DC) (DA)(DA)(DT) (DT)(DG)(DA)(DG)(DC)(DG)(DG)(DC)(DC)(DT) (DC)(DG)(DG)(DC)(DA)(DC)(DC) (DG)(DG) (DG)(DA)(DT)(DT)(DC)(DT)(DC)(DC)(DA)(DG) (DT)(DA)(DT)(DG)(DG)(DC)(DG)(DG) (DC) (DC)(DA)(DG)(DT)(DG)(DC)(DG)(DC)(DG)(DA) (DG)(DA)(DC)(DA)(DG)(DT)(DA)(DC)(DT) (DG)(DG)(DG)(DT)(DT)(DA)(DC)(DC)(DT)(DT) (DA)(DA)(DT)(DA)(DC)(DT)(DT)(DA)(DC)(DA) (DT)(DG)(DA)(DC)(DA)(DG)(DG)(DA)(DT) (DG)(DT)(DA)(DT)(DA)(DT)(DA)(DT)(DC)(DT) (DG) (DA)(DC)(DA)(DC)(DG)(DT)(DG)(DC) (DC)(DT)(DG)(DG)(DA)(DG)(DA)(DC)(DT)(DA) (DG)(DG) (DG)(DA)(DG)(DT)(DA)(DA)(DT) (DC)(DC)(DC)(DC)(DT)(DT)(DG)(DG)(DC)(DG) (DG)(DT)(DT) (DA)(DA)(DA)(DA)(DC)(DG) (DC)(DG)(DG)(DG)(DG)(DG)(DA)(DC)(DA)(DG) (DC)(DG)(DC)(DG) (DT)(DA)(DC)(DG)(DT) (DG)(DC)(DG)(DT)(DT)(DT)(DA)(DA)(DG)(DC) (DG)(DG)(DT)(DG)(DC) (DT)(DA)(DG)(DA) (DG)(DC)(DT)(DG)(DT)(DC)(DT)(DA)(DC)(DG) (DA)(DC)(DC)(DA)(DA)(DT) (DT)(DG)(DA) (DG)(DC)(DG)(DG)(DC)(DC)(DT)(DC)(DG)(DG) (DC)(DA)(DC)(DC)(DG)(DG)(DG) (DA)(DT) (DT)(DC)(DT)(DC)(DC)(DA)(DG)(DT)(DA)(DT) (DG)(DG)(DC)(DG)(DG)(DC)(DC)(DA) (DG) (DT)(DG)(DC)(DG)(DC)(DG)(DA)(DG)(DA)(DC) (DA)(DG)(DT)(DA)(DC)(DT)(DG)(DG)(DG) (DT)(DT)(DA)(DC)(DC)(DT)(DT)(DA)(DA)(DT) (DA)(DC)(DT)(DT)(DA)(DC)(DA)(DT)(DG)(DA) (DC)(DA)(DG)(DG)(DA)(DT)(DG)(DT)(DA) (DT)(DA)(DT)(DA)(DT)(DC)(DT)(DG)(DA)(DC) (DA) (DC)(DG)(DT)(DG)(DC)(DC)(DT)(DG) (DG)(DA)(DG)(DA)(DC)(DT)(DA)(DG)(DG)(DG) (DA)(DG) (DT)(DA)(DA)(DT)(DC)(DC)(DC) (DC)(DT)(DT)(DG)(DG)(DC)(DG)(DG)(DT)(DT) (DA)(DA)(DA) (DA)(DC)(DG)(DC)(DG)(DG) (DG)(DG)(DG)(DA)(DC)(DA)(DG)(DC)(DG)(DC) (DG)(DT)(DA)(DC) (DG)(DT)(DG)(DC)(DG) (DT)(DT)(DT)(DA)(DA)(DG)(DC)(DG)(DG)(DT) (DG)(DC)(DT)(DA)(DG) (DA)(DG)(DC)(DT) (DG)(DT)(DC)(DT)(DA)(DC)(DG)(DA)(DC)(DC) (DA)(DA)(DT)(DT)(DG)(DA) (DG)(DC)(DG) (DG)(DC)(DC)(DT)(DC)(DG)(DG)(DC)(DA)(DC) (DC)(DG)(DG)(DG)(DA)(DT)(DT) (DC)(DT) (DC)(DC)(DA)(DG)(DT)(DA)(DT)(DG)(DG)(DC) (DG)(DG)(DC)(DC)(DA)(DG)(DT)(DG) (DC) (DG)(DC)(DG)(DA)(DG)(DA)(DC)(DA)(DG)(DT) (DA)(DC)(DT)(DG)(DG)(DG)(DT)(DT)(DA) (DC)(DC)(DT)(DT)(DA)(DA)(DT)(DA)(DC)(DT) (DT)(DA)(DC)(DA)(DT)(DG)(DA)(DC)(DA)(DG) (DG)(DA)(DT)(DG)(DT)(DA)(DT)(DA)(DT) (DA)(DT)(DC)(DT)(DG)(DA)(DC)(DA)(DC)(DG) (DT) (DG)(DC)(DC)(DT)(DG)(DG)(DA)(DG) (DA)(DC)(DT)(DA)(DG)(DG)(DG)(DA)(DG)(DT) (DA)(DA) (DT)(DC)(DC)(DC)(DC)(DT)(DT) (DG)(DG)(DC)(DG)(DG)(DT)(DT)(DA)(DA)(DA) (DA)(DC)(DG) (DC)(DG)(DG)(DG)(DG)(DG) (DA)(DC)(DA)(DG)(DC)(DG)(DC)(DG)(DT)(DA) (DC)(DG)(DT)(DG) (DC)(DG)(DT)(DT)(DT) (DA)(DA)(DG)(DC)(DG)(DG)(DT)(DG)(DC)(DT) (DA)(DG)(DA)(DG)(DC) (DT)(DG)(DT)(DC) (DT)(DA)(DC)(DG)(DA)(DC)(DC)(DA)(DA)(DT) (DT)(DG)(DA)(DG)(DC)(DG) (DG)(DC)(DC) (DT)(DC)(DG)(DG)(DC)(DA)(DC)(DC)(DG)(DG) (DG)(DA)(DT)(DT)(DC)(DT)(DC) (DC)(DA) (DG)(DT)(DA)(DT)(DG)(DG)(DC)(DG)(DG)(DC) (DC)(DA)(DG)(DT)(DG)(DC)(DG)(DC) (DG) (DA)(DG)(DA)(DC)(DG)(DA)(DT) |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 (6k x 4k) / Average electron dose: 60.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |