 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7a01 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | The Halastavi arva virus intergenic region IRES promotes translation by the simplest possible initiation mechanism | ||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||
 キーワード キーワード | RIBOSOME / Dicistrovirus Halastavi arva virus Intergenic region Internal ribosome entry site IRES RNA pseudoknot ribosome SERBP1 CrPV IGR IRES | ||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / Translesion synthesis by REV1 / Recognition of DNA damage by PCNA-containing replication complex / Translesion Synthesis by POLH / Downregulation of ERBB4 signaling / Spry regulation of FGF signaling / Downregulation of ERBB2:ERBB3 signaling / NOD1/2 Signaling Pathway ...Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / Translesion synthesis by REV1 / Recognition of DNA damage by PCNA-containing replication complex / Translesion Synthesis by POLH / Downregulation of ERBB4 signaling / Spry regulation of FGF signaling / Downregulation of ERBB2:ERBB3 signaling / NOD1/2 Signaling Pathway / APC/C:Cdc20 mediated degradation of Cyclin B / APC-Cdc20 mediated degradation of Nek2A / EGFR downregulation / TCF dependent signaling in response to WNT / NRIF signals cell death from the nucleus / p75NTR recruits signalling complexes / NF-kB is activated and signals survival / Activated NOTCH1 Transmits Signal to the Nucleus / Downregulation of TGF-beta receptor signaling / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / Downregulation of SMAD2/3:SMAD4 transcriptional activity / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / Senescence-Associated Secretory Phenotype (SASP) / Regulation of innate immune responses to cytosolic DNA / activated TAK1 mediates p38 MAPK activation / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / Regulation of FZD by ubiquitination / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Regulation of TNFR1 signaling / TNFR1-induced NF-kappa-B signaling pathway / Translesion synthesis by POLK / Translesion synthesis by POLI / Regulation of necroptotic cell death / HDR through Homologous Recombination (HRR) / Josephin domain DUBs / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / Processing of DNA double-strand break ends / Formation of Incision Complex in GG-NER / Gap-filling DNA repair synthesis and ligation in GG-NER / Dual Incision in GG-NER / Fanconi Anemia Pathway / Regulation of TP53 Activity through Phosphorylation / Regulation of TP53 Degradation / Regulation of TP53 Activity through Methylation / Negative regulation of MET activity / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Downregulation of ERBB2 signaling / E3 ubiquitin ligases ubiquitinate target proteins / Regulation of PTEN localization / ER Quality Control Compartment (ERQC) / Regulation of expression of SLITs and ROBOs / Interferon alpha/beta signaling / Endosomal Sorting Complex Required For Transport (ESCRT) / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / IKK complex recruitment mediated by RIP1 / IRAK2 mediated activation of TAK1 complex / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Alpha-protein kinase 1 signaling pathway / RAS processing / Pexophagy / Negative regulation of FLT3 / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Regulation of NF-kappa B signaling / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / Regulation of pyruvate metabolism / SCF-beta-TrCP mediated degradation of Emi1 / Termination of translesion DNA synthesis / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Ovarian tumor domain proteases / Cyclin D associated events in G1 / Negative regulators of DDX58/IFIH1 signaling / Regulation of BACH1 activity / Negative regulation of FGFR1 signaling / Negative regulation of FGFR2 signaling / Negative regulation of FGFR3 signaling / Negative regulation of FGFR4 signaling / Negative regulation of MAPK pathway / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Inactivation of CSF3 (G-CSF) signaling / Iron uptake and transport / Deactivation of the beta-catenin transactivating complex / Metalloprotease DUBs / Formation of TC-NER Pre-Incision Complex / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / Major pathway of rRNA processing in the nucleolus and cytosol / GTP hydrolysis and joining of the 60S ribosomal subunit / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Autodegradation of the E3 ubiquitin ligase COP1 / Asymmetric localization of PCP proteins / Degradation of AXIN / Degradation of DVL / Hedgehog ligand biogenesis / Hedgehog 'on' state / TNFR2 non-canonical NF-kB pathway / DNA Damage Recognition in GG-NER / Assembly of the pre-replicative complex 類似検索 - 分子機能 | ||||||||||||||||||
| 生物種 |  Halastavi arva RNA virus (ウイルス) Halastavi arva RNA virus (ウイルス) | ||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | ||||||||||||||||||
 データ登録者 データ登録者 | Abaeva, I. / Vicens, Q. / Bochler, A. / Soufari, H. / Simonetti, A. / Pestova, T.V. / Hashem, Y. / Hellen, C.U.T. | ||||||||||||||||||
| 資金援助 |  米国, 米国,  フランス, 5件 フランス, 5件
| ||||||||||||||||||
 引用 引用 | ジャーナル: Cell Rep / 年: 2020 タイトル: The Halastavi árva Virus Intergenic Region IRES Promotes Translation by the Simplest Possible Initiation Mechanism. 著者: Irina S Abaeva / Quentin Vicens / Anthony Bochler / Heddy Soufari / Angelita Simonetti / Tatyana V Pestova / Yaser Hashem / Christopher U T Hellen / 要旨: Dicistrovirus intergenic region internal ribosomal entry sites (IGR IRESs) do not require initiator tRNA, an AUG codon, or initiation factors and jumpstart translation from the middle of the ...Dicistrovirus intergenic region internal ribosomal entry sites (IGR IRESs) do not require initiator tRNA, an AUG codon, or initiation factors and jumpstart translation from the middle of the elongation cycle via formation of IRES/80S complexes resembling the pre-translocation state. eEF2 then translocates the [codon-anticodon]-mimicking pseudoknot I (PKI) from ribosomal A sites to P sites, bringing the first sense codon into the decoding center. Halastavi árva virus (HalV) contains an IGR that is related to previously described IGR IRESs but lacks domain 2, which enables these IRESs to bind to individual 40S ribosomal subunits. By using in vitro reconstitution and cryoelectron microscopy (cryo-EM), we now report that the HalV IGR IRES functions by the simplest initiation mechanism that involves binding to 80S ribosomes such that PKI is placed in the P site, so that the A site contains the first codon that is directly accessible for decoding without prior eEF2-mediated translocation of PKI. | ||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7a01.cif.gz | 4.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7a01.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7a01.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7a01_validation.pdf.gz | 1.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7a01_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 7a01_validation.xml.gz | 347.7 KB | 表示 | |
| CIF形式データ | 7a01_validation.cif.gz | 608.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/a0/7a01ftp://data.pdbj.org/pub/pdb/validation_reports/a0/7a01 | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj



































- 集合体
集合体
| 登録構造単位 | 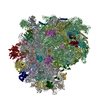
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 5種, 5分子 E1e2h2d2K3
| #1: RNA鎖 | 分子量: 48550.469 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Halastavi arva RNA virus (ウイルス)発現宿主:  |
|---|---|
| #2: RNA鎖 | 分子量: 1239030.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: RNA鎖 | 分子量: 50143.648 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #4: RNA鎖 | 分子量: 38691.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #49: RNA鎖 | 分子量: 580908.938 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-Uncharacterized ... , 10種, 10分子 p2m292J2M2W2a5B3C3H5
| #5: タンパク質 | 分子量: 8092.675 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #8: タンパク質 | 分子量: 13856.360 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 26570.105 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #28: タンパク質 | 分子量: 14319.816 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #31: タンパク質 | 分子量: 17689.461 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #37: タンパク質 | 分子量: 21627.215 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #56: タンパク質 | 分子量: 14544.659 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #68: タンパク質 | 分子量: 15975.753 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #72: タンパク質 | 分子量: 14969.445 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #78: タンパク質 | 分子量: 15812.286 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 19種, 19分子 k2q2t2u2v2x2y2A2F2H2K2U2425272U3W3E3I3
| #6: タンパク質 | 分子量: 13972.425 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #10: タンパク質 | 分子量: 16489.363 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 41101.414 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #13: タンパク質 | 分子量: 12635.694 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #14: タンパク質 | 分子量: 15022.021 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #16: タンパク質 | 分子量: 12449.612 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #17: タンパク質 | 分子量: 12994.621 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 14435.403 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #24: タンパク質 | 分子量: 12198.603 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #26: タンパク質 | 分子量: 17758.678 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #29: タンパク質 | 分子量: 21747.293 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #35: タンパク質 | 分子量: 26658.754 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #44: タンパク質 | 分子量: 24879.422 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #45: タンパク質 | 分子量: 45119.812 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #47: タンパク質 | 分子量: 18478.791 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #59: タンパク質 | 分子量: 23390.816 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #61: タンパク質 | 分子量: 24091.453 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #76: タンパク質 | 分子量: 11773.953 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #80: タンパク質 | 分子量: 11765.890 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+Ribosomal protein ... , 25種, 25分子 l2o2C2D2I2L2X2Y202128213G3G5a3A3T3Y3J5N3f3F3d3H3M3
-60S RIBOSOMAL PROTEIN ... , 13種, 13分子 r2w2B2E2G2S2T2V2223262X3a7
| #11: タンパク質 | 分子量: 8706.283 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #15: タンパク質 | 分子量: 23144.859 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #20: タンパク質 | 分子量: 11888.371 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #23: タンパク質 | 分子量: 6199.574 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #25: タンパク質 | 分子量: 34006.312 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #33: タンパク質 | 分子量: 21556.482 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #34: タンパク質 | 分子量: 22881.131 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #36: タンパク質 | 分子量: 27452.500 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #42: タンパク質 | 分子量: 15704.635 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #43: タンパク質 | 分子量: 21613.033 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #46: タンパク質 | 分子量: 20677.316 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #62: タンパク質・ペプチド | 分子量: 3213.075 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #85: タンパク質 | 分子量: 24200.525 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-40S RIBOSOMAL PROTEIN ... , 12種, 12分子 s3Q3V3j3b3c3D3e3P3I5L3O3
| #50: タンパク質・ペプチド | 分子量: 5010.986 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #52: タンパク質 | 分子量: 21431.742 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #60: タンパク質 | 分子量: 24759.145 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #64: タンパク質 | 分子量: 29527.721 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #67: タンパク質 | 分子量: 27471.535 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #71: タンパク質 | 分子量: 24003.012 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #74: タンパク質 | 分子量: 9043.276 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #75: タンパク質 | 分子量: 13766.122 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #79: タンパク質 | 分子量: 21603.229 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #81: タンパク質 | 分子量: 14604.264 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #82: タンパク質 | 分子量: 9348.990 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #84: タンパク質 | 分子量: 11184.231 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質・ペプチド / 非ポリマー , 2種, 7分子 R2
| #32: タンパク質・ペプチド | 分子量: 4063.805 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #86: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Zn |
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) | 生物種: | ||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 60 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 42135 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 94.82 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
