 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6muo: CENP-A nucleosome bound by two copies of CENP-C(CD) and one copy ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6muo | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | CENP-A nucleosome bound by two copies of CENP-C(CD) and one copy CENP-N(NT) | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | NUCLEAR PROTEIN / centromere / CENP-A / kinetochore / nucleosome | |||||||||
| Function / homology |  Function and homology information Function and homology informationspindle attachment to meiosis I kinetochore / centromeric DNA binding / CENP-A containing chromatin assembly / protein localization to chromosome, centromeric region / kinetochore assembly / attachment of mitotic spindle microtubules to kinetochore / condensed chromosome, centromeric region / inner kinetochore / establishment of mitotic spindle orientation / mitotic cytokinesis ...spindle attachment to meiosis I kinetochore / centromeric DNA binding / CENP-A containing chromatin assembly / protein localization to chromosome, centromeric region / kinetochore assembly / attachment of mitotic spindle microtubules to kinetochore / condensed chromosome, centromeric region / inner kinetochore / establishment of mitotic spindle orientation / mitotic cytokinesis / chromosome, centromeric region / pericentric heterochromatin / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Mitotic Prometaphase / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / EML4 and NUDC in mitotic spindle formation / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / Resolution of Sister Chromatid Cohesion / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / chromosome segregation / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / RHO GTPases Activate Formins / HDMs demethylate histones / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / NoRC negatively regulates rRNA expression / Negative Regulation of CDH1 Gene Transcription / G2/M DNA damage checkpoint / kinetochore / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / Meiotic recombination / Pre-NOTCH Transcription and Translation / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / RMTs methylate histone arginines / Metalloprotease DUBs / HCMV Early Events / structural constituent of chromatin / Separation of Sister Chromatids / UCH proteinases / nucleosome / mitotic cell cycle / nucleosome assembly / heterochromatin formation / HATs acetylate histones / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / chromatin organization / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / midbody / Oxidative Stress Induced Senescence / Estrogen-dependent gene expression / chromosome, telomeric region / Ub-specific processing proteases / nuclear body / Amyloid fiber formation / protein heterodimerization activity / negative regulation of cell population proliferation / cell division / chromatin binding / protein-containing complex / DNA binding / RNA binding / extracellular exosome / extracellular region / nucleoplasm / membrane / identical protein binding Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | |||||||||
 Authors Authors | Allu, P.K. / Black, B.E. | |||||||||
| Funding support |  United States, 1items United States, 1items
| |||||||||
 Citation Citation | Journal: Curr Biol / Year: 2019 Title: Structure of the Human Core Centromeric Nucleosome Complex. Authors: Praveen Kumar Allu / Jennine M Dawicki-McKenna / Trevor Van Eeuwen / Moriya Slavin / Merav Braitbard / Chen Xu / Nir Kalisman / Kenji Murakami / Ben E Black /  Abstract: Centromeric nucleosomes are at the interface of the chromosome and the kinetochore that connects to spindle microtubules in mitosis. The core centromeric nucleosome complex (CCNC) harbors the ...Centromeric nucleosomes are at the interface of the chromosome and the kinetochore that connects to spindle microtubules in mitosis. The core centromeric nucleosome complex (CCNC) harbors the histone H3 variant, CENP-A, and its binding proteins, CENP-C (through its central domain; CD) and CENP-N (through its N-terminal domain; NT). CENP-C can engage nucleosomes through two domains: the CD and the CENP-C motif (CM). CENP-C is part of the CCNC by virtue of its high specificity for CENP-A nucleosomes and ability to stabilize CENP-A at the centromere. CENP-C is thought to engage a neighboring nucleosome, either one containing conventional H3 or CENP-A, and a crystal structure of a nucleosome complex containing two copies of CENP-C was reported. Recent structures containing a single copy of CENP-N bound to the CENP-A nucleosome in the absence of CENP-C were reported. Here, we find that one copy of CENP-N is lost for every two copies of CENP-C on centromeric chromatin just prior to kinetochore formation. We present the structures of symmetric and asymmetric forms of the CCNC that vary in CENP-N stoichiometry. Our structures explain how the central domain of CENP-C achieves its high specificity for CENP-A nucleosomes and how CENP-C and CENP-N sandwich the histone H4 tail. The natural centromeric DNA path in our structures corresponds to symmetric surfaces for CCNC assembly, deviating from what is observed in prior structures using artificial sequences. At mitosis, we propose that CCNC asymmetry accommodates its asymmetric connections at the chromosome/kinetochore interface. VIDEO ABSTRACT. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6muo.cif.gz | 328.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6muo.ent.gz | 251.5 KB | Display | PDB format |
| PDBx/mmJSON format | 6muo.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/mu/6muoftp://data.pdbj.org/pub/pdb/validation_reports/mu/6muo | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 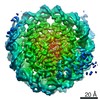 9250MC  9251C  9252C  6mupC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj













































- Assembly
Assembly
| Deposited unit | 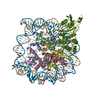
|
|---|---|
| 1 |
|
-Components
-Protein , 4 types, 8 molecules AEBFCGDH
| #1: Protein | Mass: 11993.037 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CENPAProduction host:  References: UniProt: P49450 #2: Protein | Mass: 10604.521 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, ...Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 Production host: References: UniProt: P62805 #3: Protein | Mass: 11494.393 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: HIST1H2AC, H2AFLProduction host: References: UniProt: Q93077 #4: Protein | Mass: 10249.723 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: HIST2H2BFProduction host: References: UniProt: Q5QNW6 |
|---|
-DNA/RNA (147- ... , 2 types, 2 molecules IJ
| #5: DNA chain | Mass: 45141.918 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
|---|---|
| #6: DNA chain | Mass: 45582.188 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
-Centromere protein ... , 2 types, 3 molecules KLM
| #7: Protein/peptide | Mass: 2508.834 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) / References: UniProt: Q03188#8: Protein | | Mass: 25052.885 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CENPN, C16orf60, ICEN32, BM-309Production host: References: UniProt: Q96H22 |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: CENP-A chromatin complex bound with CENP-C and CENP-N of CCAN kinetochore components Type: ORGANELLE OR CELLULAR COMPONENT / Entity ID: all / Source: MULTIPLE SOURCES |
|---|---|
| Molecular weight | Value: 0.276 MDa / Experimental value: YES |
| Source (natural) | Organism: Homo sapiens (human) |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.5 / Details: 10 mM HEPES, 50 mN NaCl, 1 mM EDTA, 1mM DTT |
| Specimen | Conc.: 0.5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Details: unspecified / Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: C-flat-1.2/1.3 4C |
| Vitrification | Instrument: LEICA EM CPC / Cryogen name: ETHANE / Details: Blot for 8 seconds before plunging |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: OTHER / Nominal magnification: 130000 X / Nominal defocus max: 3000 nm / Nominal defocus min: 500 nm / Cs: 2.7 mm / Alignment procedure: BASIC |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 40 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 QUANTUM (4k x 4k) / Num. of grids imaged: 2 |
| Image scans | Movie frames/image: 40 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 113950 / Symmetry type: POINT |
