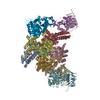
Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
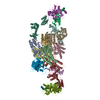
| Entry | Database: PDB / ID: 6i3m | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | eIF2B:eIF2 complex, phosphorylated on eIF2 alpha serine 52. | |||||||||||||||
 Components Components |
| |||||||||||||||
 Keywords Keywords | TRANSLATION / Translational control eIF2 phosphorylation Integrated stress response eIF2B TRANSLATION | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of cellular response to amino acid starvation / positive regulation of cellular response to heat / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / ABC-family protein mediated transport / eukaryotic translation initiation factor 2B complex / methionyl-initiator methionine tRNA binding / eukaryotic translation initiation factor 2 complex / guanyl-nucleotide exchange factor complex / multi-eIF complex ...negative regulation of cellular response to amino acid starvation / positive regulation of cellular response to heat / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / ABC-family protein mediated transport / eukaryotic translation initiation factor 2B complex / methionyl-initiator methionine tRNA binding / eukaryotic translation initiation factor 2 complex / guanyl-nucleotide exchange factor complex / multi-eIF complex / eukaryotic 43S preinitiation complex / positive regulation of cellular response to amino acid starvation / formation of translation preinitiation complex / eukaryotic 48S preinitiation complex / positive regulation of translational fidelity / protein-synthesizing GTPase / regulation of translational initiation / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / cytoplasmic translational initiation / L13a-mediated translational silencing of Ceruloplasmin expression / positive regulation of translational initiation / enzyme regulator activity / translation initiation factor activity / translation initiation factor binding / guanyl-nucleotide exchange factor activity / translational initiation / cytoplasmic stress granule / ribosome binding / ribosome / mRNA binding / GTPase activity / GTP binding / mitochondrion / RNA binding / zinc ion binding / cytoplasm / cytosol Similarity search - Function | |||||||||||||||
| Biological species |  | |||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.93 Å | |||||||||||||||
 Authors Authors | Adomavicius, T. / Roseman, A.M. / Pavitt, G.D. | |||||||||||||||
| Funding support |  United Kingdom, 4items United Kingdom, 4items
| |||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2019 Title: The structural basis of translational control by eIF2 phosphorylation. Authors: Tomas Adomavicius / Margherita Guaita / Yu Zhou / Martin D Jennings / Zakia Latif / Alan M Roseman / Graham D Pavitt /  Abstract: Protein synthesis in eukaryotes is controlled by signals and stresses via a common pathway, called the integrated stress response (ISR). Phosphorylation of the translation initiation factor eIF2 ...Protein synthesis in eukaryotes is controlled by signals and stresses via a common pathway, called the integrated stress response (ISR). Phosphorylation of the translation initiation factor eIF2 alpha at a conserved serine residue mediates translational control at the ISR core. To provide insight into the mechanism of translational control we have determined the structures of eIF2 both in phosphorylated and unphosphorylated forms bound with its nucleotide exchange factor eIF2B by electron cryomicroscopy. The structures reveal that eIF2 undergoes large rearrangements to promote binding of eIF2α to the regulatory core of eIF2B comprised of the eIF2B alpha, beta and delta subunits. Only minor differences are observed between eIF2 and eIF2αP binding to eIF2B, suggesting that the higher affinity of eIF2αP for eIF2B drives translational control. We present a model for controlled nucleotide exchange and initiator tRNA binding to the eIF2/eIF2B complex. #1: Journal: Elife / Year: 2017 Title: Fail-safe control of translation initiation by dissociation of eIF2α phosphorylated ternary complexes. Authors: Martin D Jennings / Christopher J Kershaw / Tomas Adomavicius / Graham D Pavitt / Abstract: Phosphorylation of eIF2α controls translation initiation by restricting the levels of active eIF2-GTP/Met-tRNAi ternary complexes (TC). This modulates the expression of all eukaryotic mRNAs and ...Phosphorylation of eIF2α controls translation initiation by restricting the levels of active eIF2-GTP/Met-tRNAi ternary complexes (TC). This modulates the expression of all eukaryotic mRNAs and contributes to the cellular integrated stress response. Key to controlling the activity of eIF2 are translation factors eIF2B and eIF5, thought to primarily function with eIF2-GDP and TC respectively. Using a steady-state kinetics approach with purified proteins we demonstrate that eIF2B binds to eIF2 with equal affinity irrespective of the presence or absence of competing guanine nucleotides. We show that eIF2B can compete with Met-tRNAi for eIF2-GTP and can destabilize TC. When TC is formed with unphosphorylated eIF2, eIF5 can out-compete eIF2B to stabilize TC/eIF5 complexes. However when TC/eIF5 is formed with phosphorylated eIF2, eIF2B outcompetes eIF5 and destabilizes TC. These data uncover competition between eIF2B and eIF5 for TC and identify that phosphorylated eIF2-GTP translation initiation intermediate complexes can be inhibited by eIF2B. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6i3m.cif.gz | 847.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6i3m.ent.gz | 666.7 KB | Display | PDB format |
| PDBx/mmJSON format | 6i3m.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/i3/6i3mftp://data.pdbj.org/pub/pdb/validation_reports/i3/6i3m | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4404MC  4428C  6i7tC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj








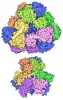






- Assembly
Assembly
| Deposited unit | 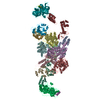
|
|---|---|
| 1 |
|
-Components
-Translation initiation factor eIF-2B subunit ... , 5 types, 10 molecules ABCDEFGHIJ
| #1: Protein | Mass: 34062.027 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #2: Protein | Mass: 70945.195 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #3: Protein | Mass: 42621.441 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #4: Protein | Mass: 81249.062 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #5: Protein | Mass: 65768.320 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|
-Eukaryotic translation initiation factor 2 subunit ... , 3 types, 6 molecules KLMNOP
| #6: Protein | Mass: 34843.633 Da / Num. of mol.: 2 / Mutation: serine 52 to SEP Source method: isolated from a genetically manipulated source Source: (gene. exp.) #7: Protein | Mass: 31631.309 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #8: Protein | Mass: 57942.699 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Complex of translation initiation factors eIF2 and eIF2B Type: COMPLEX / Entity ID: all / Source: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.838 MDa / Experimental value: YES | ||||||||||||||||||||
| Source (natural) | Organism: | ||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||
| Buffer solution | pH: 7.5 Details: Solutions for sample preparation were made fresh, filter sterilized, and degassed. | ||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||
| Specimen | Conc.: 0.25 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||
| Specimen support | Details: Ultra-thin carbon support film, 3nm - on lacey carbon grids from Agar Scientific were used for 35 degree tilted data collection. For zero tilt data collection, 200 mesh Au Quantifoil, R2/2 grids were used. Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: Homemade | ||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 294 K / Details: Blot for 2 seconds before plunging |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS / Details: Tilt at zero and 35 degrees. |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Calibrated magnification: 37313 X / Nominal defocus max: 4000 nm / Nominal defocus min: 1500 nm / Calibrated defocus min: 1000 nm / Calibrated defocus max: 5500 nm / Cs: 2.7 mm / Alignment procedure: BASIC |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 12 sec. / Electron dose: 40 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 QUANTUM (4k x 4k) / Num. of grids imaged: 2 / Num. of real images: 4533 Details: 2 separate data collections for zero degree (2255 images) and tilted (2278 images) specimen. Both on Titan Krios |
| EM imaging optics | Energyfilter name: GIF Quantum LS / Energyfilter slit width: 20 eV |
| Image scans | Sampling size: 5 µm / Width: 3838 / Height: 3710 / Movie frames/image: 48 / Used frames/image: 2-48 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: CTF determination was performed per micrograph initially. After particle picking and initial reconstruction, per particle CTF determination was performed. Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 655000 Details: Initially, approximately 3000 particles were picked manually and used for 2D classification. Then 4 best looking 2D class average images were used as references for automated particle ...Details: Initially, approximately 3000 particles were picked manually and used for 2D classification. Then 4 best looking 2D class average images were used as references for automated particle picking in RELION. For the zero tilt data collection, 406000 particles were picked. For the 35 degree tilt data collection, 249000 particles were picked | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C2 (2 fold cyclic) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.93 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 64541 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Source name: PDB / Type: experimental model
|