 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
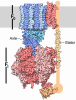
Yorodumi- PDB-5tj5: Atomic model for the membrane-embedded motor of a eukaryotic V-ATPase -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5tj5 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Atomic model for the membrane-embedded motor of a eukaryotic V-ATPase | |||||||||||||||
 Components Components | (V-type proton ATPase subunit ...) x 7 | |||||||||||||||
 Keywords Keywords | MOTOR PROTEIN / Rotary ATPase / Vacuolar-type ATPase / Electron Cryomicroscopy / Vo region / Membrane protein | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationcell wall mannoprotein biosynthetic process / ATPase-coupled ion transmembrane transporter activity / protein localization to vacuolar membrane / cellular response to alkaline pH / Insulin receptor recycling / Transferrin endocytosis and recycling / polyphosphate metabolic process / ROS and RNS production in phagocytes / Golgi lumen acidification / Amino acids regulate mTORC1 ...cell wall mannoprotein biosynthetic process / ATPase-coupled ion transmembrane transporter activity / protein localization to vacuolar membrane / cellular response to alkaline pH / Insulin receptor recycling / Transferrin endocytosis and recycling / polyphosphate metabolic process / ROS and RNS production in phagocytes / Golgi lumen acidification / Amino acids regulate mTORC1 / P-type proton-exporting transporter activity / vacuolar transport / vacuolar proton-transporting V-type ATPase, V0 domain / endosomal lumen acidification / vacuole organization / proton-transporting V-type ATPase complex / fungal-type vacuole / protein targeting to vacuole / vacuolar proton-transporting V-type ATPase complex / cellular hyperosmotic response / vacuolar acidification / fungal-type vacuole membrane / phosphatidylinositol-3,5-bisphosphate binding / proton transmembrane transporter activity / proton-transporting ATPase activity, rotational mechanism / intracellular copper ion homeostasis / Neutrophil degranulation / proton transmembrane transport / cell periphery / transmembrane transport / endocytosis / ATPase binding / protein-containing complex assembly / intracellular iron ion homeostasis / membrane raft / Golgi membrane / membrane Similarity search - Function | |||||||||||||||
| Biological species |  | |||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.9 Å | |||||||||||||||
 Authors Authors | Mazhab-Jafari, M.T. / Rohou, A. / Schmidt, C. / Bueler, S.A. / Benlekbir, S. / Robinson, C.V. / Rubinstein, J.L. | |||||||||||||||
| Funding support |  Canada, Canada,  United Kingdom, European Union, 4items United Kingdom, European Union, 4items
| |||||||||||||||
 Citation Citation | Journal: Nature / Year: 2016 Title: Atomic model for the membrane-embedded V motor of a eukaryotic V-ATPase. Authors: Mohammad T Mazhab-Jafari / Alexis Rohou / Carla Schmidt / Stephanie A Bueler / Samir Benlekbir / Carol V Robinson / John L Rubinstein /  Abstract: Vacuolar-type ATPases (V-ATPases) are ATP-powered proton pumps involved in processes such as endocytosis, lysosomal degradation, secondary transport, TOR signalling, and osteoclast and kidney ...Vacuolar-type ATPases (V-ATPases) are ATP-powered proton pumps involved in processes such as endocytosis, lysosomal degradation, secondary transport, TOR signalling, and osteoclast and kidney function. ATP hydrolysis in the soluble catalytic V region drives proton translocation through the membrane-embedded V region via rotation of a rotor subcomplex. Variability in the structure of the intact enzyme has prevented construction of an atomic model for the membrane-embedded motor of any rotary ATPase. We induced dissociation and auto-inhibition of the V and V regions of the V-ATPase by starving the yeast Saccharomyces cerevisiae, allowing us to obtain a ~3.9-Å resolution electron cryomicroscopy map of the V complex and build atomic models for the majority of its subunits. The analysis reveals the structures of subunits acc'c″de and a protein that we identify and propose to be a new subunit (subunit f). A large cavity between subunit a and the c-ring creates a cytoplasmic half-channel for protons. The c-ring has an asymmetric distribution of proton-carrying Glu residues, with the Glu residue of subunit c″ interacting with Arg735 of subunit a. The structure suggests sequential protonation and deprotonation of the c-ring, with ATP-hydrolysis-driven rotation causing protonation of a Glu residue at the cytoplasmic half-channel and subsequent deprotonation of a Glu residue at a luminal half-channel. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5tj5.cif.gz | 358.5 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5tj5.ent.gz | 265.3 KB | Display | PDB format |
| PDBx/mmJSON format | 5tj5.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/tj/5tj5ftp://data.pdbj.org/pub/pdb/validation_reports/tj/5tj5 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  8409MC  8363C  8364C  8367C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj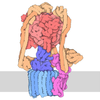










- Assembly
Assembly
| Deposited unit | 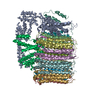
|
|---|---|
| 1 |
|
-Components
-V-type proton ATPase subunit ... , 7 types, 14 molecules ABDEFGHIJMNLOP
| #1: Protein | Mass: 70067.984 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: P32563 | ||||||
|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 22610.641 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: P23968 | ||||||
| #3: Protein | Mass: 15195.271 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: P32842 | ||||||
| #4: Protein | Mass: 15218.087 Da / Num. of mol.: 8 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: P25515 #5: Protein | | Mass: 6476.928 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: Q3E7B6 #6: Protein | | Mass: 4613.678 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c #7: Protein | | Mass: 32854.238 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) Strain: ATCC 204508 / S288c / References: UniProt: P32366 |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Vo region of the V-ATPase from Saccharomyces cerevisiae Type: COMPLEX / Entity ID: #1-#8 / Source: NATURAL |
|---|---|
| Molecular weight | Value: 0.3 MDa / Experimental value: NO |
| Source (natural) | Organism: |
| Buffer solution | pH: 7.5 Details: 50 mM Tris-HCl pH 7.5, 150 mM NaCl, 10 micro M Bafilomycin |
| Specimen | Conc.: 2.5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: The Vo region was solubilized in amphipol A8-35 (Anatrace) |
| Specimen support | Grid material: COPPER/RHODIUM / Grid mesh size: 400 divisions/in. / Grid type: Maxtaform |
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE-PROPANE / Humidity: 100 % / Chamber temperature: 277.3 K / Details: Modified for ethane/propane |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS Details: 70 micro meter objective aperture, illuminated area of 1.58 micro meter diameter |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 37000 X / Calibrated magnification: 64350 X / Calibrated defocus min: 800 nm / Calibrated defocus max: 3400 nm / Cs: 2.7 mm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 21 sec. / Electron dose: 100 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 4365 |
| Image scans | Movie frames/image: 70 |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.10.1_2155: / Classification: refinement | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||
| Image processing | Details: Images were recorded in "super-resolution" mode and then resampled by Fourier cropping. | ||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 657975 | ||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.9 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 462842 / Algorithm: FOURIER SPACE Details: Data beyond 6 A resolution were omitted from refinement. Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: OTHER / Space: REAL Details: phenix.real_space_refine REFINEMENT TARGET: REAL-SPACE (WEIGHTED MAP SUM AT ATOM CENTERS) MODEL TO MAP FIT. CC(ACROSS WHOLE MAP VOLUME): 0.6772 CC(ONLY ACROSS ATOMS IN THE MODEL): 0.6842 | ||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|
