 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4bip | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Homology model of coxsackievirus A7 (CAV7) full capsid proteins. | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | VIRUS / PICORNAVIRUS / ENTEROVIRUS / HEV-A | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報symbiont genome entry into host cell via pore formation in plasma membrane / virion component / viral capsid / host cell / host cell cytoplasm / symbiont-mediated suppression of host gene expression / symbiont entry into host cell / virion attachment to host cell / structural molecule activity 類似検索 - 分子機能 | |||||||||
| 生物種 |  HUMAN COXSACKIEVIRUS A7 (コクサッキーウイルス) HUMAN COXSACKIEVIRUS A7 (コクサッキーウイルス) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 8.23 Å | |||||||||
 データ登録者 データ登録者 | Seitsonen, J.J.T. / Shakeel, S. / Susi, P. / Pandurangan, A.P. / Sinkovits, R.S. / Hyvonen, H. / Laurinmaki, P. / Yla-Pelto, J. / Topf, M. / Hyypia, T. / Butcher, S.J. | |||||||||
 引用 引用 | ジャーナル: J Struct Biol / 年: 2014 タイトル: Combined approaches to flexible fitting and assessment in virus capsids undergoing conformational change. 著者: Arun Prasad Pandurangan / Shabih Shakeel / Sarah Jane Butcher / Maya Topf /   要旨: Fitting of atomic components into electron cryo-microscopy (cryoEM) density maps is routinely used to understand the structure and function of macromolecular machines. Many fitting methods have been ...Fitting of atomic components into electron cryo-microscopy (cryoEM) density maps is routinely used to understand the structure and function of macromolecular machines. Many fitting methods have been developed, but a standard protocol for successful fitting and assessment of fitted models has yet to be agreed upon among the experts in the field. Here, we created and tested a protocol that highlights important issues related to homology modelling, density map segmentation, rigid and flexible fitting, as well as the assessment of fits. As part of it, we use two different flexible fitting methods (Flex-EM and iMODfit) and demonstrate how combining the analysis of multiple fits and model assessment could result in an improved model. The protocol is applied to the case of the mature and empty capsids of Coxsackievirus A7 (CAV7) by flexibly fitting homology models into the corresponding cryoEM density maps at 8.2 and 6.1Å resolution. As a result, and due to the improved homology models (derived from recently solved crystal structures of a close homolog - EV71 capsid - in mature and empty forms), the final models present an improvement over previously published models. In close agreement with the capsid expansion observed in the EV71 structures, the new CAV7 models reveal that the expansion is accompanied by ∼5° counterclockwise rotation of the asymmetric unit, predominantly contributed by the capsid protein VP1. The protocol could be applied not only to viral capsids but also to many other complexes characterised by a combination of atomic structure modelling and cryoEM density fitting. #1: ジャーナル: J Virol / 年: 2012タイトル: Structural analysis of coxsackievirus A7 reveals conformational changes associated with uncoating. 著者: Jani J T Seitsonen / Shabih Shakeel / Petri Susi / Arun P Pandurangan / Robert S Sinkovits / Heini Hyvönen / Pasi Laurinmäki / Jani Ylä-Pelto / Maya Topf / Timo Hyypiä / Sarah J Butcher / 要旨: Coxsackievirus A7 (CAV7) is a rarely detected and poorly characterized serotype of the Enterovirus species Human enterovirus A (HEV-A) within the Picornaviridae family. The CAV7-USSR strain has ...Coxsackievirus A7 (CAV7) is a rarely detected and poorly characterized serotype of the Enterovirus species Human enterovirus A (HEV-A) within the Picornaviridae family. The CAV7-USSR strain has caused polio-like epidemics and was originally thought to represent the fourth poliovirus type, but later evidence linked this strain to the CAV7-Parker prototype. Another isolate, CAV7-275/58, was also serologically similar to Parker but was noninfectious in a mouse model. Sequencing of the genomic region encoding the capsid proteins of the USSR and 275/58 strains and subsequent comparison with the corresponding amino acid sequences of the Parker strain revealed that the Parker and USSR strains are nearly identical, while the 275/58 strain is more distant. Using electron cryomicroscopy and three-dimensional image reconstruction, the structures of the CAV7-USSR virion and empty capsid were resolved to 8.2-Å and 6.1-Å resolutions, respectively. This is one of the first detailed structural analyses of the HEV-A species. Using homology modeling, reconstruction segmentation, and flexible fitting, we constructed a pseudoatomic T = 1 (pseudo T = 3) model incorporating the three major capsid proteins (VP1 to VP3), addressed the conformational changes of the capsid and its constituent viral proteins occurring during RNA release, and mapped the capsid proteins' variable regions to the structure. During uncoating, VP4 and RNA are released analogously to poliovirus 1, the interfaces of VP2 and VP3 are rearranged, and VP1 rotates. Variable regions in the capsid proteins were predicted to map mainly to the surface of VP1 and are thus likely to affect the tropism and pathogenicity of CAV7. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
 UCSF Chimera
UCSF Chimera- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4bip.cif.gz | 96.2 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4bip.ent.gz | 55.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 4bip.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/bi/4bipftp://data.pdbj.org/pub/pdb/validation_reports/bi/4bip | HTTPS FTP |
|---|
-関連構造データ






-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 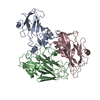
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | x 60
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | x 5
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | x 6
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: (シェーンフリース記号: I (正20面体型対称)) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 非結晶学的対称性 (NCS) | NCS oper:
|