 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3j47 | ||||||
|---|---|---|---|---|---|---|---|
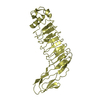
| タイトル | Formation of an intricate helical bundle dictates the assembly of the 26S proteasome lid | ||||||
 要素 要素 | (26S proteasome regulatory subunit ...) x 8 | ||||||
 キーワード キーワード | PROTEIN BINDING / alpha helix bundle / hybrid method / flexible fitting | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報: / proteasome storage granule assembly / peroxisome fission / protein deneddylation / COP9 signalosome / proteasome regulatory particle / mitochondrial fission / proteasome regulatory particle, lid subcomplex / metal-dependent deubiquitinase activity / Proteasome assembly ...: / proteasome storage granule assembly / peroxisome fission / protein deneddylation / COP9 signalosome / proteasome regulatory particle / mitochondrial fission / proteasome regulatory particle, lid subcomplex / metal-dependent deubiquitinase activity / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / KEAP1-NFE2L2 pathway / Neddylation / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Orc1 removal from chromatin / MAPK6/MAPK4 signaling / proteasome binding / regulation of protein catabolic process / proteasome storage granule / Antigen processing: Ubiquitination & Proteasome degradation / protein deubiquitination / Ub-specific processing proteases / proteasome assembly / enzyme regulator activity / Neutrophil degranulation / proteasome complex / metallopeptidase activity / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / ubiquitin-dependent protein catabolic process / proteasome-mediated ubiquitin-dependent protein catabolic process / ubiquitinyl hydrolase 1 / cysteine-type deubiquitinase activity / structural molecule activity / mitochondrion / metal ion binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 7.4 Å | ||||||
 データ登録者 データ登録者 | Estrin, E. / Lopez-Blanco, J.R. / Chacon, P. / Martin, A. | ||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2012 タイトル: Near-atomic resolution structural model of the yeast 26S proteasome. 著者: Florian Beck / Pia Unverdorben / Stefan Bohn / Andreas Schweitzer / Günter Pfeifer / Eri Sakata / Stephan Nickell / Jürgen M Plitzko / Elizabeth Villa / Wolfgang Baumeister / Friedrich Förster /  要旨: The 26S proteasome operates at the executive end of the ubiquitin-proteasome pathway. Here, we present a cryo-EM structure of the Saccharomyces cerevisiae 26S proteasome at a resolution of 7.4 Å or ...The 26S proteasome operates at the executive end of the ubiquitin-proteasome pathway. Here, we present a cryo-EM structure of the Saccharomyces cerevisiae 26S proteasome at a resolution of 7.4 Å or 6.7 Å (Fourier-Shell Correlation of 0.5 or 0.3, respectively). We used this map in conjunction with molecular dynamics-based flexible fitting to build a near-atomic resolution model of the holocomplex. The quality of the map allowed us to assign α-helices, the predominant secondary structure element of the regulatory particle subunits, throughout the entire map. We were able to determine the architecture of the Rpn8/Rpn11 heterodimer, which had hitherto remained elusive. The MPN domain of Rpn11 is positioned directly above the AAA-ATPase N-ring suggesting that Rpn11 deubiquitylates substrates immediately following commitment and prior to their unfolding by the AAA-ATPase module. The MPN domain of Rpn11 dimerizes with that of Rpn8 and the C-termini of both subunits form long helices, which are integral parts of a coiled-coil module. Together with the C-terminal helices of the six PCI-domain subunits they form a very large coiled-coil bundle, which appears to serve as a flexible anchoring device for all the lid subunits. | ||||||
| 履歴 |
| ||||||
| Remark 0 | THIS ENTRY 3J47 CONTAINS A STRUCTURAL MODEL FIT TO AN ELECTRON MICROSCOPY MAP (EMD-2165) DETERMINED ...THIS ENTRY 3J47 CONTAINS A STRUCTURAL MODEL FIT TO AN ELECTRON MICROSCOPY MAP (EMD-2165) DETERMINED ORIGINALLY BY AUTHORS: F.BECK, P.UNVERDORBEN, S.BOHN, A.SCHWEITZER, G.PFEIFER, E.SAKATA, S.NICKELL, J.M.PLITZKO, E.VILLA, W.BAUMEISTER, F.FORSTER |
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j47.cif.gz | 68.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j47.ent.gz | 47.9 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 3j47.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j4/3j47ftp://data.pdbj.org/pub/pdb/validation_reports/j4/3j47 | HTTPS FTP |
|---|
-関連構造データ
| 関連構造データ |  2165M M: このデータのモデリングに利用したマップデータ |
|---|---|
| 類似構造データ |
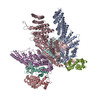



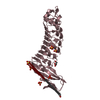

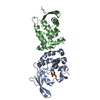



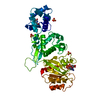

-リンク
 PDBj
PDBj













- 集合体
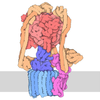
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-26S proteasome regulatory subunit ... , 8種, 8分子 VUOPQRST
| #1: タンパク質 | 分子量: 8013.857 Da / 分子数: 1 断片: last three C-terminal helices (UNP residues 230-298) 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 13965.031 Da / 分子数: 1 断片: last three C-terminal helices (UNP residues 188-308) 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質・ペプチド | 分子量: 3353.889 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 360-387) / 由来タイプ: 天然 / 由来: (天然) |
| #4: タンパク質・ペプチド | 分子量: 3984.469 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 409-442) / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質・ペプチド | 分子量: 2741.054 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 407-431) / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質・ペプチド | 分子量: 2890.363 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 397-422) / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質・ペプチド | 分子量: 2924.156 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 455-478) / 由来タイプ: 天然 / 由来: (天然) |
| #8: タンパク質・ペプチド | 分子量: 1955.234 Da / 分子数: 1 / 断片: C-terminal helix (UNP residues 256-272) / 由来タイプ: 天然 / 由来: (天然) |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: 26S proteasome / タイプ: COMPLEX |
|---|---|
| 緩衝液 | pH: 7.1 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: HOMEMADE PLUNGER / 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2012年3月15日 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 150000 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 1500 nm |
| 撮影 | 電子線照射量: 25 e/Å2 フィルム・検出器のモデル: TVIPS TEMCAM-F816 (8k x 8k) |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 解像度: 7.4 Å / 解像度の算出法: FSC 0.5 CUT-OFF / 粒子像の数: 2464694 / 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL / Target criteria: Cross-correlation coefficient 詳細: METHOD--Hybrid method + flexible fitting REFINEMENT PROTOCOL--Hybrid method DETAILS--Initial model was done with an in house hybrid method (EMTEGRATOR) that integrates topology constraints ...詳細: METHOD--Hybrid method + flexible fitting REFINEMENT PROTOCOL--Hybrid method DETAILS--Initial model was done with an in house hybrid method (EMTEGRATOR) that integrates topology constraints with EM-map derived constraints. iMODFIT was then used for final flexible fitting. | ||||||||||||
| 精密化ステップ | サイクル: LAST
|
