regulation of ubiquitin-protein transferase activity / epithelial cell maturation involved in prostate gland development / Intra-Golgi traffic / branching involved in prostate gland morphogenesis / intra-Golgi vesicle-mediated transport / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / regulation of DNA damage checkpoint / cellular response to chemical stress ...regulation of ubiquitin-protein transferase activity / epithelial cell maturation involved in prostate gland development / Intra-Golgi traffic / branching involved in prostate gland morphogenesis / intra-Golgi vesicle-mediated transport / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / regulation of DNA damage checkpoint / cellular response to chemical stress / Cul7-RING ubiquitin ligase complex / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / target-directed miRNA degradation / elongin complex / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / VCB complex / regulation of extrinsic apoptotic signaling pathway via death domain receptors / positive regulation of protein autoubiquitination / death receptor binding / protein neddylation / NEDD8 ligase activity / Cul5-RING ubiquitin ligase complex / negative regulation of response to oxidative stress / ubiquitin-ubiquitin ligase activity / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / Cul4A-RING E3 ubiquitin ligase complex / negative regulation of type I interferon production / Cul4B-RING E3 ubiquitin ligase complex / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Cul3-RING ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / Prolactin receptor signaling / protein monoubiquitination / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / cullin family protein binding / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / Tat-mediated elongation of the HIV-1 transcript / ubiquitin-like ligase-substrate adaptor activity / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / protein K48-linked ubiquitination / RNA Polymerase II Transcription Elongation / Nuclear events stimulated by ALK signaling in cancer / Formation of RNA Pol II elongation complex / RNA Polymerase II Pre-transcription Events / positive regulation of TORC1 signaling / post-translational protein modification / regulation of cellular response to insulin stimulus / intrinsic apoptotic signaling pathway / Regulation of BACH1 activity / transcription corepressor binding / T cell activation / TP53 Regulates Transcription of DNA Repair Genes / transcription initiation at RNA polymerase II promoter / Degradation of DVL / transcription elongation by RNA polymerase II / Recognition of DNA damage by PCNA-containing replication complex / Degradation of GLI1 by the proteasome / cellular response to amino acid stimulus / Negative regulation of NOTCH4 signaling / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Vif-mediated degradation of APOBEC3G / Hedgehog 'on' state / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / DNA Damage Recognition in GG-NER / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / RING-type E3 ubiquitin transferase / negative regulation of canonical Wnt signaling pathway / Degradation of beta-catenin by the destruction complex / Inactivation of CSF3 (G-CSF) signaling / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / Formation of TC-NER Pre-Incision Complex / Dual Incision in GG-NER / Evasion by RSV of host interferon responses / NOTCH1 Intracellular Domain Regulates Transcription / Constitutive Signaling by NOTCH1 PEST Domain Mutants / Constitutive Signaling by NOTCH1 HD+PEST Domain Mutants / Regulation of expression of SLITs and ROBOs / Formation of Incision Complex in GG-NER / Dual incision in TC-NER / Interleukin-1 signaling / Gap-filling DNA repair synthesis and ligation in TC-NER / Orc1 removal from chromatin / protein polyubiquitination / Regulation of RAS by GAPs / ubiquitin-protein transferase activity / G1/S transition of mitotic cell cycle / positive regulation of protein catabolic process / Regulation of RUNX2 expression and activity / cellular response to UV / MAPK cascade / ubiquitin protein ligase activity / KEAP1-NFE2L2 pathway / Antigen processing: Ubiquitination & Proteasome degradation / positive regulation of proteasomal ubiquitin-dependent protein catabolic process 類似検索 - 分子機能
National Natural Science Foundation of China (NSFC)
中国
引用
ジャーナル: Nat Commun / 年: 2024 タイトル: Mechanism of Ψ-Pro/C-degron recognition by the CRL2 ubiquitin ligase. 著者: Xinyan Chen / Anat Raiff / Shanshan Li / Qiong Guo / Jiahai Zhang / Hualin Zhou / Richard T Timms / Xuebiao Yao / Stephen J Elledge / Itay Koren / Kaiming Zhang / Chao Xu / 要旨: The E3 ligase-degron interaction determines the specificity of the ubiquitin‒proteasome system. We recently discovered that FEM1B, a substrate receptor of Cullin 2-RING ligase (CRL2), recognizes C- ...The E3 ligase-degron interaction determines the specificity of the ubiquitin‒proteasome system. We recently discovered that FEM1B, a substrate receptor of Cullin 2-RING ligase (CRL2), recognizes C-degrons containing a C-terminal proline. By solving several cryo-EM structures of CRL2 bound to different C-degrons, we elucidate the dimeric assembly of the complex. Furthermore, we reveal distinct dimerization states of unmodified and neddylated CRL2 to uncover the NEDD8-mediated activation mechanism of CRL2. Our research also indicates that, FEM1B utilizes a bipartite mechanism to recognize both the C-terminal proline and an upstream aromatic residue within the substrate. These structural findings, complemented by in vitro ubiquitination and in vivo cell-based assays, demonstrate that CRL2-mediated polyubiquitination and subsequent protein turnover depend on both FEM1B-degron interactions and the dimerization state of the E3 ligase complex. Overall, this study deepens our molecular understanding of how Cullin-RING E3 ligase substrate selection mediates protein turnover.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 中国, 1件
中国, 1件  引用
引用


 構造の表示
構造の表示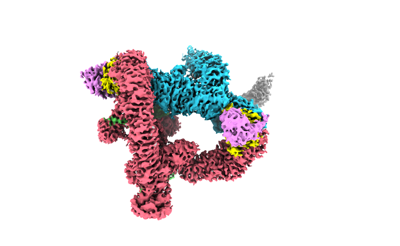
 ダウンロードとリンク
ダウンロードとリンク emd_37743.png
emd_37743.png http://ftp.pdbj.org/pub/emdb/structures/EMD-37743
http://ftp.pdbj.org/pub/emdb/structures/EMD-37743




























 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)
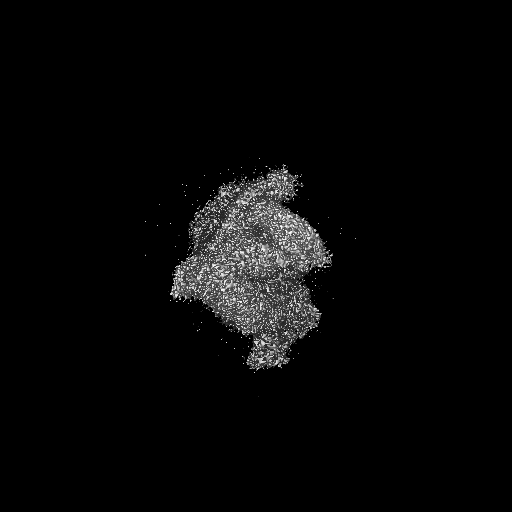
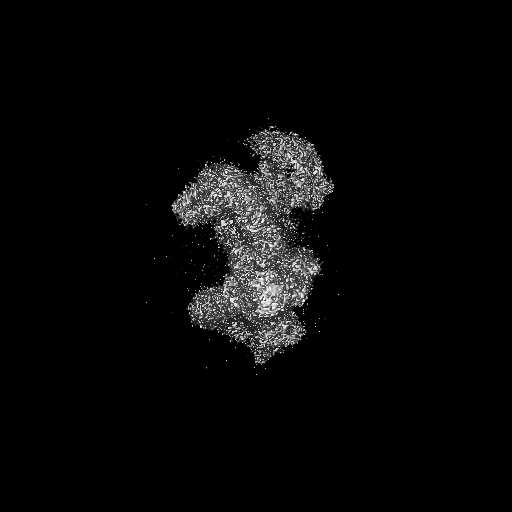
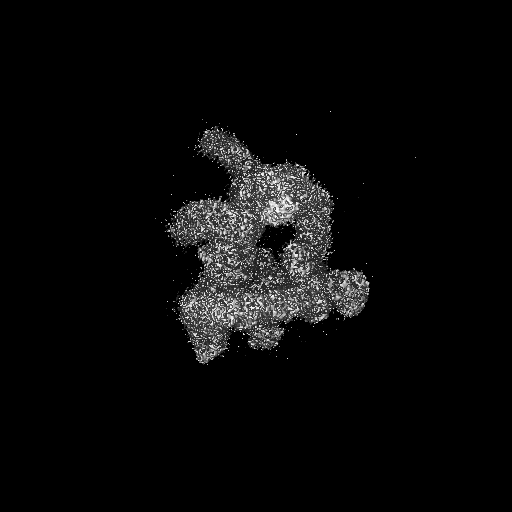









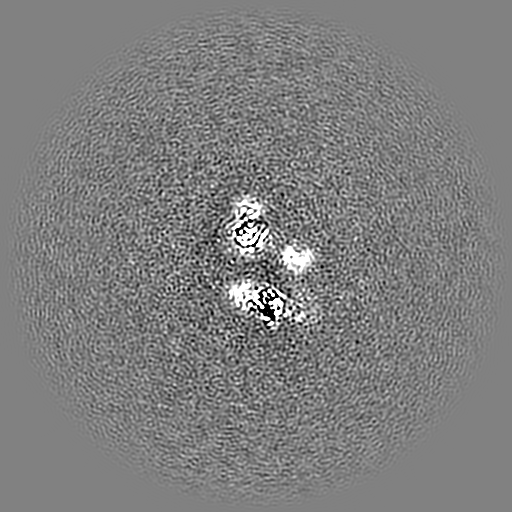

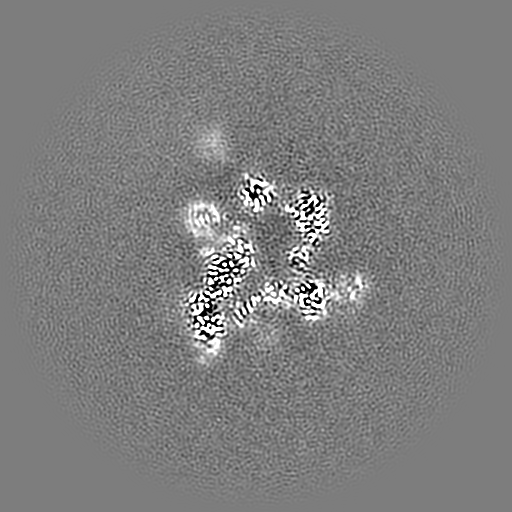





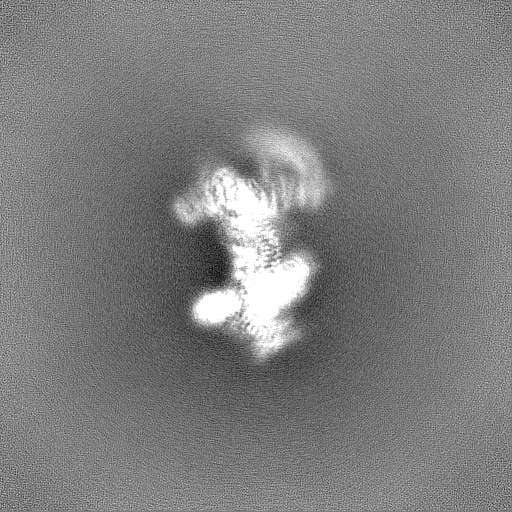
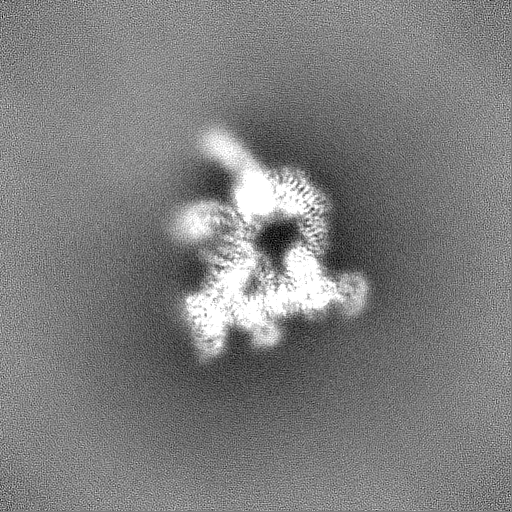


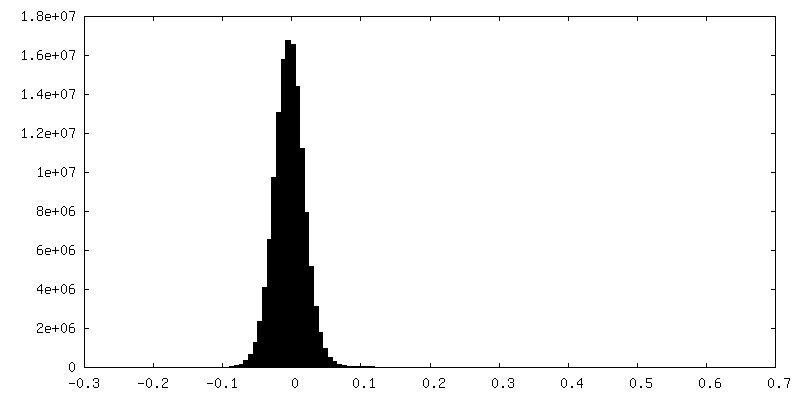



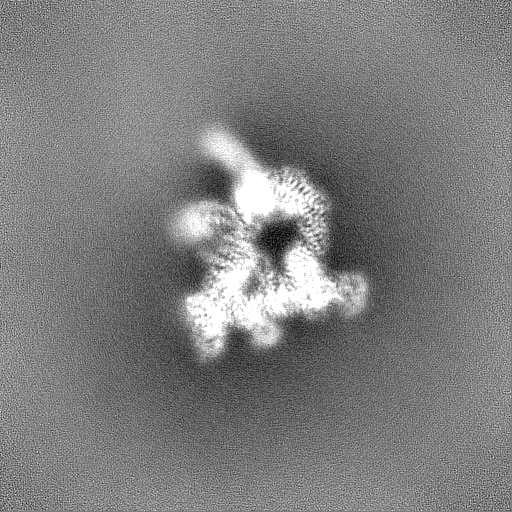


 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
