 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8wqa: Cryo-EM structure of CUL2-RBX1-ELOB-ELOC-FEM1B bound with the C-d... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8wqa | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of CUL2-RBX1-ELOB-ELOC-FEM1B bound with the C-degron of CCDC89 (conformation 1) | ||||||
 Components Components |
| ||||||
 Keywords Keywords | LIGASE / E3 ubiquitin ligase / Pro/C-degron | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of ubiquitin-protein transferase activity / epithelial cell maturation involved in prostate gland development / branching involved in prostate gland morphogenesis / negative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / target-directed miRNA degradation ...regulation of ubiquitin-protein transferase activity / epithelial cell maturation involved in prostate gland development / branching involved in prostate gland morphogenesis / negative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / target-directed miRNA degradation / regulation of DNA damage checkpoint / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / elongin complex / regulation of extrinsic apoptotic signaling pathway via death domain receptors / death receptor binding / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / protein neddylation / NEDD8 ligase activity / protein K27-linked ubiquitination / negative regulation of response to oxidative stress / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-ubiquitin ligase activity / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Cul4A-RING E3 ubiquitin ligase complex / Prolactin receptor signaling / negative regulation of mitophagy / Cul4-RING E3 ubiquitin ligase complex / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / cullin family protein binding / protein monoubiquitination / Tat-mediated elongation of the HIV-1 transcript / ubiquitin ligase complex / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / ubiquitin-like ligase-substrate adaptor activity / RNA Polymerase II Transcription Elongation / site of DNA damage / Formation of RNA Pol II elongation complex / signal transduction in response to DNA damage / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / transcription-coupled nucleotide-excision repair / RNA Polymerase II Pre-transcription Events / negative regulation of insulin receptor signaling pathway / regulation of cellular response to insulin stimulus / positive regulation of TORC1 signaling / post-translational protein modification / intrinsic apoptotic signaling pathway / T cell activation / transcription corepressor binding / Regulation of BACH1 activity / negative regulation of canonical NF-kappaB signal transduction / TP53 Regulates Transcription of DNA Repair Genes / transcription initiation at RNA polymerase II promoter / cellular response to amino acid stimulus / transcription elongation by RNA polymerase II / Degradation of DVL / G1/S transition of mitotic cell cycle / Degradation of CRY and PER proteins / negative regulation of canonical Wnt signaling pathway / Degradation of GLI1 by the proteasome / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Recognition of DNA damage by PCNA-containing replication complex / Negative regulation of NOTCH4 signaling / RING-type E3 ubiquitin transferase / Hedgehog 'on' state / Vif-mediated degradation of APOBEC3G / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Inactivation of CSF3 (G-CSF) signaling / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Evasion by RSV of host interferon responses / NOTCH1 Intracellular Domain Regulates Transcription / Degradation of beta-catenin by the destruction complex / DNA Damage Recognition in GG-NER / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Constitutive Signaling by NOTCH1 PEST Domain Mutants / Constitutive Signaling by NOTCH1 HD+PEST Domain Mutants / Dual Incision in GG-NER / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / Formation of TC-NER Pre-Incision Complex / Regulation of expression of SLITs and ROBOs / Formation of Incision Complex in GG-NER / Interleukin-1 signaling / Orc1 removal from chromatin / protein polyubiquitination / Dual incision in TC-NER Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.39 Å | ||||||
 Authors Authors | Chen, X. / Zhang, K. / Xu, C. | ||||||
| Funding support |  China, 1items China, 1items
| ||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: Mechanism of Ψ-Pro/C-degron recognition by the CRL2 ubiquitin ligase. Authors: Xinyan Chen / Anat Raiff / Shanshan Li / Qiong Guo / Jiahai Zhang / Hualin Zhou / Richard T Timms / Xuebiao Yao / Stephen J Elledge / Itay Koren / Kaiming Zhang / Chao Xu /    Abstract: The E3 ligase-degron interaction determines the specificity of the ubiquitin‒proteasome system. We recently discovered that FEM1B, a substrate receptor of Cullin 2-RING ligase (CRL2), recognizes C- ...The E3 ligase-degron interaction determines the specificity of the ubiquitin‒proteasome system. We recently discovered that FEM1B, a substrate receptor of Cullin 2-RING ligase (CRL2), recognizes C-degrons containing a C-terminal proline. By solving several cryo-EM structures of CRL2 bound to different C-degrons, we elucidate the dimeric assembly of the complex. Furthermore, we reveal distinct dimerization states of unmodified and neddylated CRL2 to uncover the NEDD8-mediated activation mechanism of CRL2. Our research also indicates that, FEM1B utilizes a bipartite mechanism to recognize both the C-terminal proline and an upstream aromatic residue within the substrate. These structural findings, complemented by in vitro ubiquitination and in vivo cell-based assays, demonstrate that CRL2-mediated polyubiquitination and subsequent protein turnover depend on both FEM1B-degron interactions and the dimerization state of the E3 ligase complex. Overall, this study deepens our molecular understanding of how Cullin-RING E3 ligase substrate selection mediates protein turnover. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8wqa.cif.gz | 639.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8wqa.ent.gz | 504.5 KB | Display | PDB format |
| PDBx/mmJSON format | 8wqa.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/wq/8wqaftp://data.pdbj.org/pub/pdb/validation_reports/wq/8wqa | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  37736MC  8wqbC 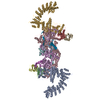 8wqcC  8wqdC  8wqeC  8wqfC 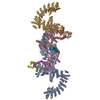 8wqgC  8wqhC  8wqiC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |








-Links
 PDBj
PDBj













- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 10 molecules CADBEGFHJI
| #1: Protein | Mass: 87114.750 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CUL2 / Production host:  #2: Protein | Mass: 70355.062 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: FEM1B, F1AA, KIAA0396 / Production host: #3: Protein | Mass: 10843.420 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ELOC, TCEB1 / Production host: #4: Protein | Mass: 13147.781 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ELOB, TCEB2 / Production host: #5: Protein | Mass: 11196.830 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RBX1, RNF75, ROC1 / Production host: |
|---|
-Protein/peptide / Non-polymers , 2 types, 8 molecules KL
| #6: Protein/peptide | Mass: 3223.647 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CCDC89, BOIP / Production host: #7: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn |
|---|
-Details
| Has ligand of interest | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Cryo-EM structure of CUL2-RBX1-ELOB-ELOC-FEM1B bound with the C-degron of CCDC89 (conformation 1) Type: COMPLEX / Entity ID: #1-#6 / Source: MULTIPLE SOURCES |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Conc.: 5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: COPPER / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R2/1 |
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277.15 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2900 nm / Nominal defocus min: 1500 nm |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Electron dose: 57.6 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Num. of real images: 4472 |
| EM imaging optics | Energyfilter name: GIF Bioquantum / Energyfilter slit width: 20 eV |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.39 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 23621 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
