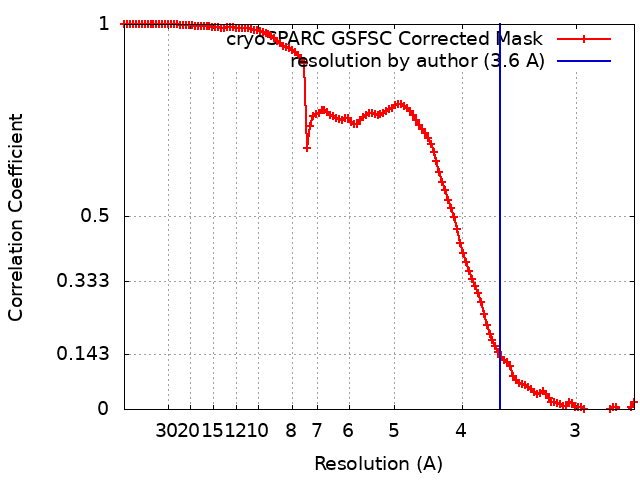
- EMDB-27811: Symmetry expansion of yeast cytoplasmic dynein-1 bound to Lis1 in... -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: EMDB / ID: EMD-27811
タイトル
Symmetry expansion of yeast cytoplasmic dynein-1 bound to Lis1 in the chi conformation.
マップデータ
試料
複合体: cytoplasmic dynein(E2448Q) bound to Lis1 in chi conformation
タンパク質・ペプチド: Dynein heavy chain, cytoplasmic
タンパク質・ペプチド: Nuclear distribution protein PAC1
リガンド: ADENOSINE-5'-TRIPHOSPHATE
リガンド: ADENOSINE-5'-DIPHOSPHATE
キーワード
dynein / motor protein / transport
機能・相同性
機能・相同性情報
positive regulation of microtubule plus-end binding / microtubule sliding / microtubule organizing center organization / nuclear migration along microtubule / vesicle transport along microtubule / minus-end-directed microtubule motor activity / dynein light intermediate chain binding / microtubule plus-end binding / cytoplasmic dynein complex / retrograde axonal transport ...positive regulation of microtubule plus-end binding / microtubule sliding / microtubule organizing center organization / nuclear migration along microtubule / vesicle transport along microtubule / minus-end-directed microtubule motor activity / dynein light intermediate chain binding / microtubule plus-end binding / cytoplasmic dynein complex / retrograde axonal transport / nuclear migration / microtubule associated complex / dynein intermediate chain binding / dynein complex binding / cytoplasmic microtubule / establishment of mitotic spindle orientation / Antigen processing: Ubiquitination & Proteasome degradation / cytoplasmic microtubule organization / mitotic spindle organization / kinetochore / spindle pole / nuclear envelope / cell cortex / cell division / ATP hydrolysis activity / ATP binding / identical protein binding / nucleus / cytoplasm 類似検索 - 分子機能
Dynein regulator LIS1 / LIS1, N-terminal / : / DYN1, AAA+ ATPase lid domain / Dynein heavy chain 3, AAA+ lid domain / AAA+ lid domain / P-loop containing dynein motor region / Dynein heavy chain, tail / Dynein heavy chain, N-terminal region 1 / Dynein heavy chain ...Dynein regulator LIS1 / LIS1, N-terminal / : / DYN1, AAA+ ATPase lid domain / Dynein heavy chain 3, AAA+ lid domain / AAA+ lid domain / P-loop containing dynein motor region / Dynein heavy chain, tail / Dynein heavy chain, N-terminal region 1 / Dynein heavy chain / Dynein heavy chain region D6 P-loop domain / Dynein heavy chain, linker / Dynein heavy chain, AAA module D4 / Dynein heavy chain, coiled coil stalk / Dynein heavy chain, hydrolytic ATP-binding dynein motor region / Dynein heavy chain, ATP-binding dynein motor region / Dynein heavy chain AAA lid domain / Dynein heavy chain AAA lid domain superfamily / Dynein heavy chain, domain 2, N-terminal / Dynein heavy chain, linker, subdomain 3 / Dynein heavy chain, AAA1 domain, small subdomain / Dynein heavy chain region D6 P-loop domain / Dynein heavy chain, N-terminal region 2 / Hydrolytic ATP binding site of dynein motor region / Microtubule-binding stalk of dynein motor / P-loop containing dynein motor region D4 / ATP-binding dynein motor region / Dynein heavy chain AAA lid domain / G-protein beta WD-40 repeat / WD40 repeat, conserved site / Trp-Asp (WD) repeats signature. / Trp-Asp (WD) repeats profile. / Trp-Asp (WD) repeats circular profile. / WD domain, G-beta repeat / WD40 repeats / WD40 repeat / WD40-repeat-containing domain superfamily / WD40/YVTN repeat-like-containing domain superfamily / ATPases associated with a variety of cellular activities / AAA+ ATPase domain / P-loop containing nucleoside triphosphate hydrolase 類似検索 - ドメイン・相同性
Dynein heavy chain, cytoplasmic / Nuclear distribution protein PAC1 類似検索 - 構成要素
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
R01 GM107214
米国
引用
ジャーナル: Nat Struct Mol Biol / 年: 2023 タイトル: Lis1 relieves cytoplasmic dynein-1 autoinhibition by acting as a molecular wedge. 著者: Eva P Karasmanis / Janice M Reimer / Agnieszka A Kendrick / Kendrick H V Nguyen / Jennifer A Rodriguez / Joey B Truong / Indrajit Lahiri / Samara L Reck-Peterson / Andres E Leschziner / 要旨: Cytoplasmic dynein-1 transports intracellular cargo towards microtubule minus ends. Dynein is autoinhibited and undergoes conformational changes to form an active complex that consists of one or two ...Cytoplasmic dynein-1 transports intracellular cargo towards microtubule minus ends. Dynein is autoinhibited and undergoes conformational changes to form an active complex that consists of one or two dynein dimers, the dynactin complex, and activating adapter(s). The Lissencephaly 1 gene, LIS1, is genetically linked to the dynein pathway from fungi to mammals and is mutated in people with the neurodevelopmental disease lissencephaly. Lis1 is required for active dynein complexes to form, but how it enables this is unclear. Here, we present a structure of two yeast dynein motor domains with two Lis1 dimers wedged in-between. The contact sites between dynein and Lis1 in this structure, termed 'Chi,' are required for Lis1's regulation of dynein in Saccharomyces cerevisiae in vivo and the formation of active human dynein-dynactin-activating adapter complexes in vitro. We propose that this structure represents an intermediate in dynein's activation pathway, revealing how Lis1 relieves dynein's autoinhibited state.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 米国, 2件
米国, 2件  引用
引用
 構造の表示
構造の表示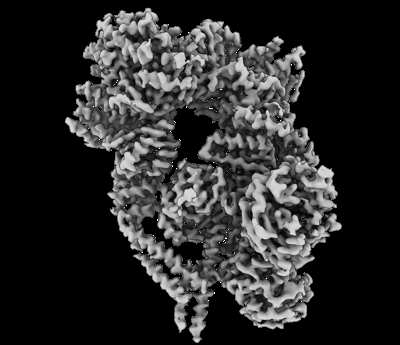
 ダウンロードとリンク
ダウンロードとリンク emd_27811.png
emd_27811.png http://ftp.pdbj.org/pub/emdb/structures/EMD-27811
http://ftp.pdbj.org/pub/emdb/structures/EMD-27811







 Z
Z Y
Y X
X
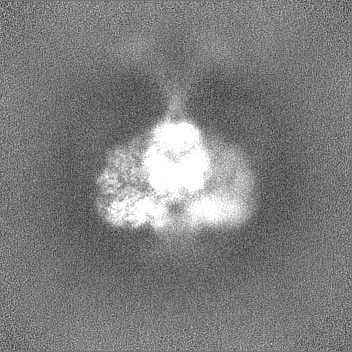
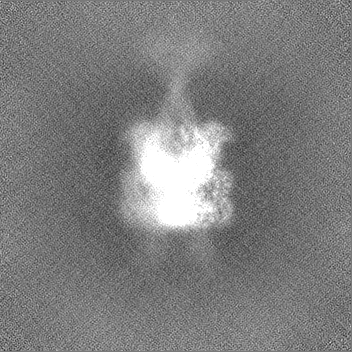

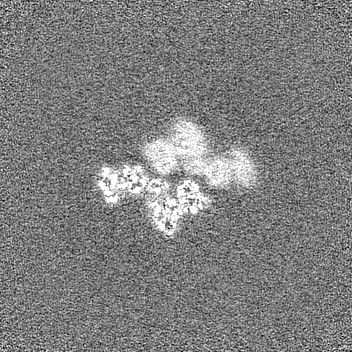
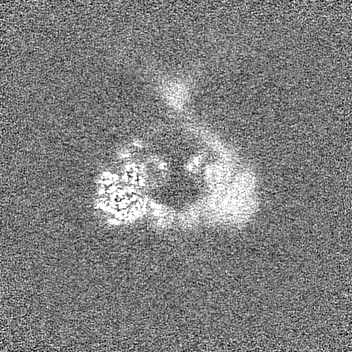
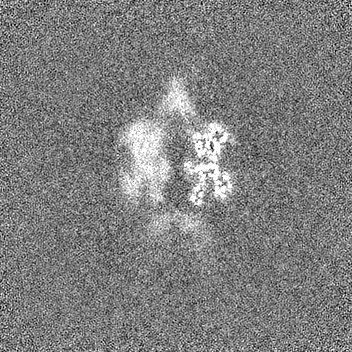


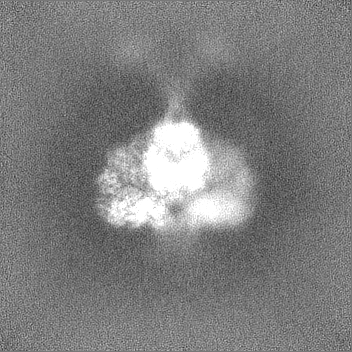

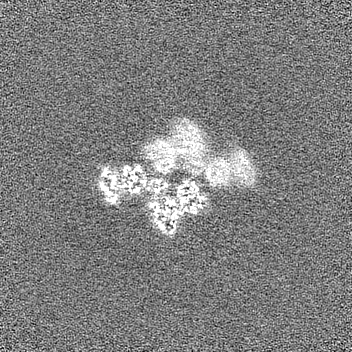
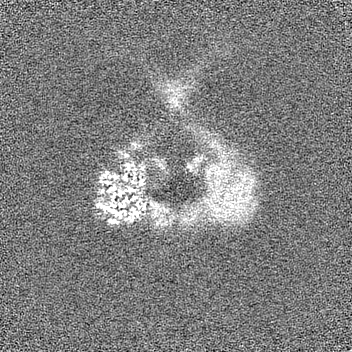

 試料の構成要素
試料の構成要素

 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN