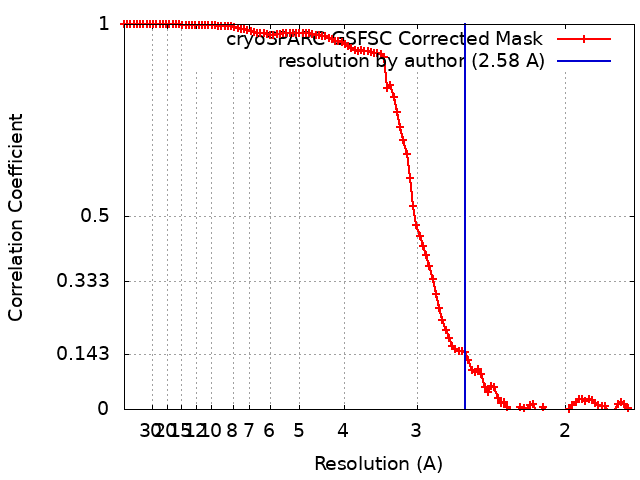
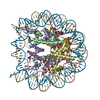
- EMDB-17740: Cryo-EM structure of NR5A2-nucleosome complex SHL+5.5 -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: EMDB / ID: EMD-17740
タイトル
Cryo-EM structure of NR5A2-nucleosome complex SHL+5.5
マップデータ
試料
複合体: Cryo-EM structure of the nucleosome containing NR5A2 motif at SHL+5.5
タンパク質・ペプチド: Histone H3.3
タンパク質・ペプチド: Histone H4
タンパク質・ペプチド: Histone H2A
タンパク質・ペプチド: Histone H2B type 1-C/E/G
DNA: DNA
DNA: DNA
タンパク質・ペプチド: Nuclear receptor subfamily 5 group A member 2
リガンド: ZINC ION
キーワード
Nucleosome / nuclear receptor / NR5A2 / DNA BINDING PROTEIN
機能・相同性
機能・相同性情報
Inhibition of DNA recombination at telomere / Deposition of new CENPA-containing nucleosomes at the centromere / SUMOylation of chromatin organization proteins / DNA Damage/Telomere Stress Induced Senescence / E3 ubiquitin ligases ubiquitinate target proteins / Regulation of gene expression in early pancreatic precursor cells / G2/M DNA damage checkpoint / Recognition and association of DNA glycosylase with site containing an affected purine / HDMs demethylate histones / Cleavage of the damaged purine ...Inhibition of DNA recombination at telomere / Deposition of new CENPA-containing nucleosomes at the centromere / SUMOylation of chromatin organization proteins / DNA Damage/Telomere Stress Induced Senescence / E3 ubiquitin ligases ubiquitinate target proteins / Regulation of gene expression in early pancreatic precursor cells / G2/M DNA damage checkpoint / Recognition and association of DNA glycosylase with site containing an affected purine / HDMs demethylate histones / Cleavage of the damaged purine / Nonhomologous End-Joining (NHEJ) / Condensation of Prophase Chromosomes / HDACs deacetylate histones / pancreas morphogenesis / PRC2 methylates histones and DNA / Processing of DNA double-strand break ends / HATs acetylate histones / calcineurin-mediated signaling / PKMTs methylate histone lysines / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / acinar cell differentiation / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / RMTs methylate histone arginines / negative regulation of chromosome condensation / tissue development / Barr body / regulation of centromere complex assembly / Factors involved in megakaryocyte development and platelet production / Estrogen-dependent gene expression / muscle cell differentiation / pericentric heterochromatin formation / inner kinetochore / bile acid metabolic process / embryo development ending in birth or egg hatching / oocyte maturation / oogenesis / homeostatic process / nucleus organization / chromosome, centromeric region / spermatid development / subtelomeric heterochromatin formation / single fertilization / RNA polymerase II core promoter sequence-specific DNA binding / protein localization to CENP-A containing chromatin / positive regulation of viral genome replication / CENP-A containing nucleosome / nucleosomal DNA binding / embryo implantation / hormone-mediated signaling pathway / innate immune response in mucosa / cellular response to leukemia inhibitory factor / transcription coregulator binding / cholesterol homeostasis / SUMOylation of intracellular receptors / multicellular organism growth / phospholipid binding / Nuclear Receptor transcription pathway / osteoblast differentiation / RNA polymerase II transcription regulator complex / structural constituent of chromatin / antimicrobial humoral immune response mediated by antimicrobial peptide / nuclear receptor activity / male gonad development / nucleosome / sequence-specific double-stranded DNA binding / nucleosome assembly / chromatin organization / chromosome / regulation of cell population proliferation / antibacterial humoral response / positive regulation of cell growth / spermatogenesis / DNA-binding transcription activator activity, RNA polymerase II-specific / Estrogen-dependent gene expression / cell population proliferation / sequence-specific DNA binding / chromosome, telomeric region / transcription cis-regulatory region binding / DNA-binding transcription factor activity, RNA polymerase II-specific / defense response to Gram-positive bacterium / protein heterodimerization activity / RNA polymerase II cis-regulatory region sequence-specific DNA binding / DNA-binding transcription factor activity / chromatin binding / regulation of DNA-templated transcription / chromatin / regulation of transcription by RNA polymerase II / positive regulation of DNA-templated transcription / positive regulation of transcription by RNA polymerase II / DNA binding / extracellular space / zinc ion binding / nucleoplasm / identical protein binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能
ジャーナル: Nat Struct Mol Biol / 年: 2024 タイトル: Nucleosome-bound NR5A2 structure reveals pioneer factor mechanism by DNA minor groove anchor competition. 著者: Wataru Kobayashi / Anna H Sappler / Daniel Bollschweiler / Maximilian Kümmecke / Jérôme Basquin / Eda Nur Arslantas / Siwat Ruangroengkulrith / Renate Hornberger / Karl Duderstadt / Kikuë Tachibana / 要旨: Gene expression during natural and induced reprogramming is controlled by pioneer transcription factors that initiate transcription from closed chromatin. Nr5a2 is a key pioneer factor that regulates ...Gene expression during natural and induced reprogramming is controlled by pioneer transcription factors that initiate transcription from closed chromatin. Nr5a2 is a key pioneer factor that regulates zygotic genome activation in totipotent embryos, pluripotency in embryonic stem cells and metabolism in adult tissues, but the mechanism of its pioneer activity remains poorly understood. Here, we present a cryo-electron microscopy structure of human NR5A2 bound to a nucleosome. The structure shows that the conserved carboxy-terminal extension (CTE) loop of the NR5A2 DNA-binding domain competes with a DNA minor groove anchor of the nucleosome and releases entry-exit site DNA. Mutational analysis showed that NR5A2 D159 of the CTE is dispensable for DNA binding but required for stable nucleosome association and persistent DNA 'unwrapping'. These findings suggest that NR5A2 belongs to an emerging class of pioneer factors that can use DNA minor groove anchor competition to destabilize nucleosomes and facilitate gene expression during reprogramming.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 Homo sapiens (ヒト) / synthetic construct (人工物)
Homo sapiens (ヒト) / synthetic construct (人工物) データ登録者
データ登録者 ドイツ, European Union, 2件
ドイツ, European Union, 2件  引用
引用 構造の表示
構造の表示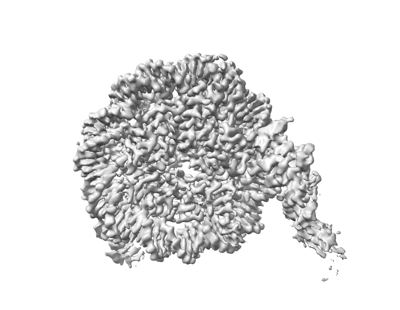
 ダウンロードとリンク
ダウンロードとリンク emd_17740.png
emd_17740.png http://ftp.pdbj.org/pub/emdb/structures/EMD-17740
http://ftp.pdbj.org/pub/emdb/structures/EMD-17740








 Z
Z Y
Y X
X



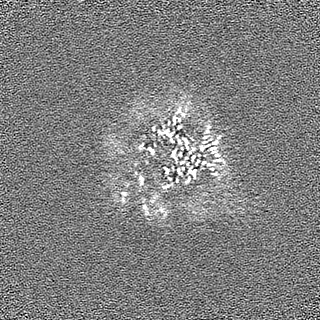




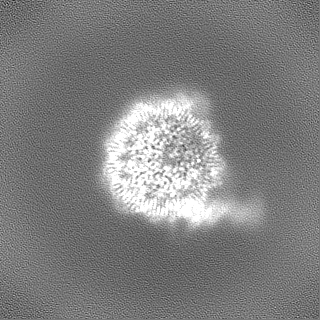
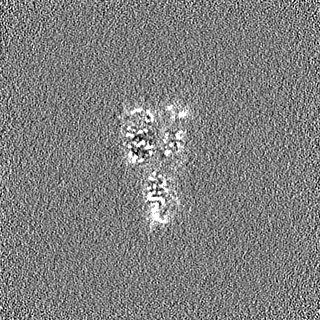

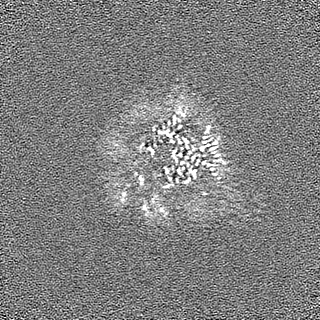
 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN