National Natural Science Foundation of China (NSFC)
31991161
China
National Natural Science Foundation of China (NSFC)
32230020
China
National Natural Science Foundation of China (NSFC)
32241029
China
National Natural Science Foundation of China (NSFC)
31730023
China
National Natural Science Foundation of China (NSFC)
31500606
China
National Natural Science Foundation of China (NSFC)
62131004
China
Chinese Academy of Sciences
XDB37010100
China
Howard Hughes Medical Institute (HHMI)
55008737
United States
Chinese Academy of Sciences
153311KYSB20170020
China
Citation
Journal: Cell Res / Year: 2024 Title: Structural basis for linker histone H5-nucleosome binding and chromatin fiber compaction. Authors: Wenyan Li / Jie Hu / Feng Song / Juan Yu / Xin Peng / Shuming Zhang / Lin Wang / Mingli Hu / Jia-Cheng Liu / Yu Wei / Xue Xiao / Yan Li / Dongyu Li / Hui Wang / Bing-Rui Zhou / Linchang Dai ...Authors: Wenyan Li / Jie Hu / Feng Song / Juan Yu / Xin Peng / Shuming Zhang / Lin Wang / Mingli Hu / Jia-Cheng Liu / Yu Wei / Xue Xiao / Yan Li / Dongyu Li / Hui Wang / Bing-Rui Zhou / Linchang Dai / Zongjun Mou / Min Zhou / Haonan Zhang / Zheng Zhou / Huidong Zhang / Yawen Bai / Jin-Qiu Zhou / Wei Li / Guohong Li / Ping Zhu / Abstract: The hierarchical packaging of chromatin fibers plays a critical role in gene regulation. The 30-nm chromatin fibers, a central-level structure bridging nucleosomal arrays to higher-order ...The hierarchical packaging of chromatin fibers plays a critical role in gene regulation. The 30-nm chromatin fibers, a central-level structure bridging nucleosomal arrays to higher-order organizations, function as the first level of transcriptional dormant chromatin. The dynamics of 30-nm chromatin fiber play a crucial role in biological processes related to DNA. Here, we report a 3.6-angstrom resolution cryogenic electron microscopy structure of H5-bound dodecanucleosome, i.e., the chromatin fiber reconstituted in the presence of linker histone H5, which shows a two-start left-handed double helical structure twisted by tetranucleosomal units. An atomic structural model of the H5-bound chromatin fiber, including an intact chromatosome, is built, which provides structural details of the full-length linker histone H5, including its N-terminal domain and an HMG-motif-like C-terminal domain. The chromatosome structure shows that H5 binds the nucleosome off-dyad through a three-contact mode in the chromatin fiber. More importantly, the H5-chromatin structure provides a fine molecular basis for the intra-tetranucleosomal and inter-tetranucleosomal interactions. In addition, we systematically validated the physiological functions and structural characteristics of the tetranucleosomal unit through a series of genetic and genomic studies in Saccharomyces cerevisiae and in vitro biophysical experiments. Furthermore, our structure reveals that multiple structural asymmetries of histone tails confer a polarity to the chromatin fiber. These findings provide structural and mechanistic insights into how a nucleosomal array folds into a higher-order chromatin fiber with a polarity in vitro and in vivo.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
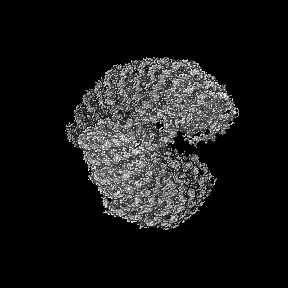
Basic information
 Map data
Map data Sample
Sample Keywords
Keywords Function and homology information
Function and homology information
 Authors
Authors China,
China,  United States, 10 items
United States, 10 items  Citation
Citation Structure visualization
Structure visualization
 Downloads & links
Downloads & links emd_38407.png
emd_38407.png http://ftp.pdbj.org/pub/emdb/structures/EMD-38407
http://ftp.pdbj.org/pub/emdb/structures/EMD-38407




 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)




















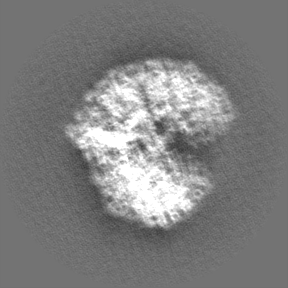



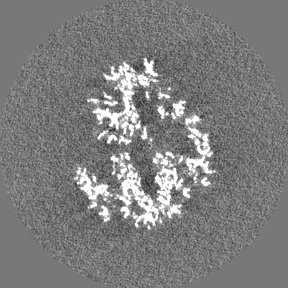









 Sample components
Sample components
 Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN

